Siitepölyn peittämä kimalainen sinihibiskuksen kukassa. – I, Luc Viatour [GFDL (http://www.gnu.org/copyleft/fdl.html), CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/) tai CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5)], lähde: Wikimedia Commons.Uusi kasvien maailmanvalloittajaryhmä syntyi, kun sammalten, liekomaisten ja saniaisten maaelämään sopeutumien lisäksi kehittyi joukko muita ominaisuuksia. Merkittävintä niistä oli siitepölyn ja siementen kehittyminen. Niihin liittyen siemenkasveille ovat ominaista myös surkastuneet gametofyytit, erilaisitiöisyys ja siemenaiheet. Näiden innovaatioiden myötä kasvien kuivankesto ja ultraviolettivalon sieto lisääntyivät, eikä hedelmöitykseen enää tarvittu vettä, joten suvullinen lisääntyminen tuli mahdolliseksi monenlaisissa ympäristöissä.
Kertauksen vuoksi: Sammalilla elinkiertoa hallitsee sukusoluja tuottava gametofyyttivaihe. Liekomaisilla ja saniaisilla vallitseva vaihe on itiöitä tuottava sporofyytti. Gametofyyttivaiheen surkastuminen jatkui siinä kehityslinjassa, jonka tulosta siemenkasvit ovat. Saniaismaisilla gametofyytin voi nähdä paljain silmin, siemenkasveilla ei voi.
Siemenkasvien mikroskooppisen pienet gametofyytit voivat kehittyä itiöistä itiöpesäkkeen sisällä, jolloin ne ovat suojassa ympäristön aiheuttamalta stressiltä, kuten kuivumiselta ja säteilyltä. Lisäksi kehittyvät gametofyytit saavat ravintoa sporofyytiltä eikä niiden tarvitse kehittyä itseyhteyttäviksi.
Suurin osa siemenettömistä kasveista on yhtäläisitiöisiä, so. ne tuottavat vain yhdenlaisia itiöitä, joista itää kaksineuvoisia gametofyyttejä. Jossain vaiheessa siemenkasvien kantamuodot kehittyivät erilaisitiöisiksi. Eräät lehdet muuntuivat isoitiöpesäkekannattimiksi, joissa on isoitiöpesäkkeitä, jotka tuottavat isoitiöitä. Niistä kehittyy naarasgametofyyttejä, jotka sisältävät munasolun. Vastaavasti pikkuitiölehtiin kasvaa pikkuitiöpesäkkeitä, jotka tuottavat pikkuitiöitä ja niistä kehittyviä koirasgametofyyttejä siittiöineen.
Kussakin isoitiöpesäkkeesä on yksi isoitiö; pikkuitiöpesäkkeissä on paljon pikkuitiöitä. Isoitiötä ja sen pesäkettä nimitetään siemenkasveilla siemenaiheeksi ja pikkuitiöpesäkettä siitepölyhiukkaseksi.
Paljassiemenisen (vas.) ja koppisiemenisen (oik.) siemenaihe. Paljassiemenisillä on yksi integumentti, jossa on siitinaukko eli mikropyle. Isoitiöpesäkkeen emosolu eli megasporosyytti on kummallakin nukelluksen sisällä. Koppisiemenisillä siemenaihe sijaitsee kukan emilehden sisällä, ja se on erilaistunut kuvassa näkyviksi vartaloksi (style) ja sikiäimeksi (ovary). Siemenaihe on sikiäimen seinämästä kasvavan kannattimen (funiculus) päässä ja siinä on kaksi integumenttia. – By derivative work: Qef (talk)Ovule-Gymno-Angio-fr.svg:Tameeria at en.wikipedia. Translation and vectorisation by Cehagenmerak. (Ovule-Gymno-Angio-fr.svg) [Public domain], from Wikimedia Commons.Megasporangio eli isoitiöpesäke pysyy emokasvin sisällä. Emokasvi kehittä megasporangion ympärille solukkoa suojaksi; näin syntyvät siemenaiheen kalvot eli integumentit (paljassiemenisillä on yksi, koppisiemenisillä kaksi integumenttia). Näin on syntynyt isoitiön, sen pesäkkeen ja intgumenttien muodostama rakenne: siemenaihe. Siinä naarasgametofyytti kehittyy isoitiöstä ja tuottaa yhden tai useampia munasoluja.
Pikkuitiöpesäkeestä kehittyy siitepölyhiukkanen. Se koostuu koirasgametofyytistä ja hiukkasen seinämästä. Seinämän uloin kerros muodostuu sporofyytin solujen erittämistä molekyyleistä. Sporopolleniiniseinämä suojaa siitepölyhiukkasta sen kulkeutuessa tuulen tai jonkin eläimen mukana siemenaiheen luo. Tapahtumaa nimitetään pölyttämiseksi (pollinaatio). Kun siitepölyhiukkanen itää, se kasvattaa siiteputken, jota myöten siittiöt pääsevät siemenaiheen sisällä olevaan naarasgametofyyttiin. Siittiöt eivät tavallisesti ole siemenkasveilla siimallisia (saniaisilla ovat).
Kortteet ja saniaiset muodostavat monilofyyttien ryhmän (Monilophyta), joka aloitti evoluutiotaivalluksensa devonikaudella noin 400 miljoonaa vuotta sitten. Kivihiilikauden (360-285 milj. v. sitten) rämeillä kortteet ja saniaiset kasvoivat yhdessä liekomaisiin kuuluvien jättiläispuiden kanssa. Nykyään saniaisia on noin 12000 lajia, ja niitä kasvaa runsaiten ja monimuotoisimmin tropiikissa, mutta myös lauhkean vyöhykkeen metsissä, ja jotkut lajit ovat sopeutuneet kuiviin olosuhteisiin.
Monilofyytit ovat läheisempää sukua siemenkasveille kuin liekomaisille. Siihen viittaa se, että siemenkasveilla ja saniaisilla on yhteisiä ominaisuuksia, joita ei löydy liekomaisilta. Saniaisten ja siemenkasvien lehdet ovat ns. megafyllejä, eli niissä on kuljetussolukkojen muodostamien suonten verkosto (liekomaisten mikrofylleissä on vain yksi lehtisuoni). Saniaiset ja siemenkasvit muodostavat kladin, evoluutiopuun oksan, jota nimitetään eufyllofyyteiksi (Euphyllophyta, ’aitolehtikasvit’). Eufyllofyyttien juuretkin ovat edistyneempää tyyppiä: ne voivat haarautua mistä kohdasta tahansa ja siten lisätä vettä ja ravinteita absorboivaa pinta-alaa tehokkaammin kuin halkihaaraisesti kasvavat liekomaisten juuret.
Monilofyyttien ryhmä sisältää seuraavat kladit (monofyleettiset ryhmät):
Equisetales: kortekasvit.
Ophioglossales: käärmeenkielikasvit.
Psilotales: haarusanikkaiskasvit.
Marattiales.
Leptosporangiaatit
Osmundales: kuningassaniaiskasvit.
Hymenophyllales: sammalsaniaiskasvit.
Gleicheniales: mm. aurinkosaniaiskasvit.
Schizaeales.
Salviniales: ormiokasvit ja kellussaniaiskasvit.
Cyatheales: saniaispuukasvit.
Polypodiales: mm. tesmasaniaiskasvit, sanikkakasvit, raunioiskasvit ja kallioimarrekasvit.
Kortekasvit
Kortteet (Equisetum) on noin 20-lajinen suku, joka muodostaa yksinään lahkon Equisetales ja kortekasvien heimon Equisetaceae. Samaan lahkoon yhdistettiin aiemmin joukko fossiilisia taksoneita, kuten jättimäiset kortepuut (Calamites). Nyt kuitenkin ollaan sitä mieltä, että nykyiset kortteet ovat läheisempää sukua saniaisille, kun taas suurin osa muinaisista fossiileista muodosti oman kehityslinjansa, jolle on annettu nimeksi Sphenophyta. Niiden rinnalla kasvoi toki jo aitoja kortteitakin, joiden fossiileja on devoni- ja kivihiilikautisissa kerrostumissa.
Kortteet on helppo tunnistaa nivelikkäistä ja lehdettömistä varsista ja niiden päähän kehittyvistä itiötähkistä. Suku on levinnyt enimmäkseen pohjoisen pallonpuoliskon lauhkeaan vyöhykkeeseen ja Etelä-Amerikkaan. Vanhan maailman tropiikissa esiintyy vain yksi laji.
Suomessa on kahdeksan alkuperäistä ja vakiintunutta kortelajia: peltokorte (Equisetum arvense), järvikorte (E. fluviatile), kangaskorte (E. hyemale), suokorte (E. palustre), lehtokorte (E. pratense), hentokorte (E. scirpoides), metsäkorte (E.sylvaticum), kirjokorte (E. variegatum).
Peltokorte kasvattaa itiöpesäketähkän ja yhteyttävän verson eriaikaisesti. Se on meillä yleinen rikkakasvi. – By Edward Step [Public domain], via Wikimedia Commons.
käärmeenkielikasvit
Käärmeenkielikasvit ( heimo Ophioglossaceae lahkossa Ophioglossales) on niin ikään lahkonsa ainoa heimo. Molekyylikello osoittaa, että ryhmä on noin 160 miljoonaa vuotta vanha. Aiemmin ei käärmeenkielikasveja pidetty saniaisten sukulaisina pääasiassa siksi, että kehittyvät lehdet eivät ole kiertyneet sisäänpäin kuten muilla saniaisilla.
Käärmeenkielikasveja tavataan lähes kaikkialla maapallolla kuivimpia alueita lukuun ottamatta. Sukuja on neljä ja lajeja noin 80; maassamme kasvaa niistä kahdeksan: noidanlukkoja (Botrychium) ja käärmeenkieliä (Ophioglossum).
Ketonoidanlukko (Botrychium lunaria) kasvaa Suomessa yleisenä hiekkaisilla kedoilla. Lehti on erilaistunut yhteyttäväksi osaksi ja itiöpesäkelehdeksi. – Tekijä: 511 Botrychium vulgare, B. matricariae.jpg: Dim Grits (511 Botrychium vulgare, B. matricariae.jpg) [Public domain], lähde: Wikimedia Commons.
haarusanikkaiskasvit
Haarusanikkaisia (heimo Psilotaceae) pidettiin varhemmin varhaisimpien halkihaaraisesti kasvavien putkilokasvien (esim. Rhynia, Psilophyton) sukulaisina, mutta samankaltaisuus on osoittautunut konvergenttisen evoluution tulokseksi.
Lehtien yksinkertaisuus muistuttaa liekomaisten mikrofyllejä. Haarusanikkaiset ovat kaukaista sukua käärmeenkielikasveille, ja niillä on joitakin morfologisia yhtäläisyyksiä.
Haarusanikkaiskasveja tunnetaan noin 12 lajia, jotka ryhmitellään kahdeksi suvuksi. Lajit kasvavat kaikkialla tropiikissa ja lämpimillä alueilla, missä on tarpeeksi kosteutta. Meitä lähinnä niitä on Lounais-Euroopassa.
Tässäkin lahkossa on vain yksi heimo, Marattiaceae, joka käsittää kuusi sukua ja noin 135 lajia. Lahkoa edustavat myös eräät fossiiliset puumaiset saniaiset. Marattiaceae-heimon saniaiset kasvavat maapollon trooppisilla ja subtrooppisilla alueilla; Euroopasta ne puuttuvat.
Marattia salicina on suurikokoinen saniainen, joka kasvaa luontaisesti Uudessa-Seelannissa ja Etelämeren saarilla. – Tekijä: Kahuroa [Public domain], Wikimedia Commonsista.
leptosporangiaatit
Jäljellä olevat seitsemän saniaislahkoa muodostavat yhtenäisen kladin eli haaran evoluutiopuussa. Niille on ominaista se, että itiöpesäkkeet kehittyvät yhdestä ainoasta lehden päällysketon solusta, kun sen sijaan edellisillä saniaislahkoilla, liekomaisilla ja siemenkasveilla itiöpesäkkeet saavat alkunsa monesta solusta – näitä sanotaan eusporangiaateiksi. Suurin osa nykysaniaisista kuuluu leptosporangiaatteihin.
Leptosporangiaateilla itiöpesäkeryhmät eli sorukset ovat usein suomumaisen rakenteen, indusiumin, peittämiä. Indusiumin muoto auttaa usein näiden saniaisten tunnistamisessa.
kuningassaniaiskasvit
Osmundales-lahkon ainoa heimo on Osmundaceae, kuningassaniaiskasvit. Niiden sukulaisia tunnetaan jo permikauden fossiileista. Nykyisessä heimossa on neljä sukua ja 23 lajia, jotka ovat levinneet lähes koko maapallolle kuumia autiomaita lukuun ottamatta. Kuningassaniaiskasvien lahkossa esiintyy itiöpesäketyyppi, joka on käärmeenkielikasvien ja leptosporangiaattien välimuoto.
Lahkossa Hymenophyllales on ainoastaan sammalsaniaiskasvien heimo (Hymenophyllaceae). Saniaisille epätyypillisesti sen lajeilla on vain yhden solukerroksen paksuiset, ohuet lehdet. Lahko erilaistui noin 243 miljoonaa vuotta sitten.
Sammalsaniaiskasveihin kuuluu kaksi sukua ja noin 650 lajia. Niitä tavataan kaikkialta tropiikissa ja lauhkean vyöhykkeen sademetsissä, lähimmät esiintymät ovat Norjassa.
Lahkossa Gleicheniales on kolme heimoa, jotka muodostavat yhtenäisen ryhmän, ja joilla on pitkä fossiilihistoria. Ne ovat hyviä menneisyyden – ja nykyisyyden – trooppisten kasvillisuusalueiden indikaattoreita. Nykyiset heimot ovat:
Aurinkosaniaiskasvit, Gleicheniales: sukuja 6, lajeja n. 165; heimo on pantrooppinen (kasvaa kaikkialla tropiikissa). Gleichenia polypodioides -aurinkosaniainen Etelä-Afrikasta. – Tekijä: Paul venter [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0) tai GFDL (http://www.gnu.org/copyleft/fdl.html)], Wikimedia Commonsista
Dipteridaceae: sukuja 2, lajeja 9; trooppinen Aasia ja Australaasia.
Matoniaceae: sukuja 2, lajeja 4; Malesian kasvistoalue.
Schizaeales
Lahkolla ja sen ainoalla heimolla Schizaeaceae ei ole suomenkielistä nimeä, eikä ihme, sillä se on levinneisyydeltään trooppinen, vain Amerikassa se ulottuu lauhkeaan vyöhykkeeseen. Sukuja siinä on neljä, lajeja noin 190. Erikoispiirteenä on lehtien reunoissa olevat sporangioforit eli pesäkekannattimet, joissa itiöpesäkkeet sijaitsevat.
Schizaea pusilla Pohjois-Amerikasta. – Tekijä: Nathaniel Lord Britton & Addison Brown [Public domain], lähde: Wikimedia Commons.
ormio- ja kellussaniaiskasvit
Nämä kaksi vesisaniaisheimoa muodostavat lahkon Salviniales. Sen lajit ovat erilaisitiöisiä. Itiöpesäkkeet sijaitsevat ns. sporokarpeissa, jotka ovat kovapintaisia rakenteita. Tämän arvellaan olevan sopeutuma vesielämään.
Ormiokasvit, Marsileaceae: sukuja 3, lajeja n. 65; levinneisyydeltään lähes yleismaailmallinen kylmimpiä ja kuivimpia alueita lukuun ottamatta; Suomessa ormio (Pilularia globulifera) kirkasvetisissä järvissä mineraalipohjalla Etelä-Hämeessä harvinaisena.
Ormio. – Wikimedia commons. Original book source: Prof. Dr. Otto Wilhelm Thomé Flora von Deutschland, Österreich und der Schweiz 1885, Gera, Germany.
Kellussaniaiskasvit, Salviniaceae: vapaasti kelluvia vesikasveja; sukuja 2, lajeja n. 20; eteläinen Pohjois-Amerikka, Etelä-Amerikka, Etelä-Eurooppa ja Länsi-Aasia sekä Vanhan maailman tropiikki. Kellussaniainen (Salvinia natans). – Tekijä: Le.Loup.Gris [GFDL 1.3 (www.gnu.org/licenses/fdl-1.3.html) tai CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], Wikimedia Commonsista.
saniaispuukasvit
Saniaspuukasvit, Cyatheaceae, ovat ainoa heimo lahkossaan. Aiemmin ne jaettiin kahdeksaksi eri heimoksi, sillä ryhmässä on hyvin vaihtelevannäköisiä lajeja; on mm. puumaisia ja maavarrellisia, tavanomaisemman saniaismaisia. Kaikilla on kuitenkin samanlainen itiöpesäkkeen rakenne.
Saniaispuukasvit jaetaan kahdeksaksi alaheimoksi, joissa on yhteensä 12 sukua ja noin 700 lajia. Ne kasvavat tropiikissa ja eteläisellä pallonpuoliskolla myös subtrooppisilla ja kosteilla lauhkeilla alueilla.
Jäljellä olevat kahdeksan saniaisheimoa muodostavat lahkon Polypodiales. Siihen kuuluu nykyisten saniaislajien suuri enemmistö, kuten myös Suomen saniaisten enemmistö.
Lonchitidaceae: sukuja 1, lajeja 2; trooppinen Amerikka, trooppinen Afrikka ja Madagaskar.
Saccolomataceae: sukuja 1, lajeja 12; trooppinen Amerikka, Madagaskar, trooppinen Aasia, Australaasia ja läntiset Tyynenmeren saaret.
Cystodiaceae: sukuja 1, lajeja 1; Borneo ja siitä itään olevat saaret.
Lindsaeaceae: sukuja 6, lajeja 220; tropiikki.
Tesmasaniaiskasvit, Dennstaedtiaceae: sukuja 10, lajeja 240; yleismaailmallinen levinneisyys, puuttuu kylmyys- ja kuivuusaavikoilta; Suomessa sananjalka (Pteridium aquilinum), joka on levinnyt lähes kaikkialle planeetallamme. Sananjalka. – Tekijä: by Thomas Moore ; edited by John Lindley ; nature-printed by Henry Bradbury. (The ferns of Great Britain and Ireland.) [Public domain], lähde: Wikimedia Commons.
Sanikkakasvit, Pteridaceae: sukuja n. 45, lajeja n. 1150; lähes kosmopoliittinen heimo. Suomessa harvinisena pohjoisessa liesu (Cryptogramma crispa). Liesu. – Tekijä: Bernd Haynold [GFDL (http://www.gnu.org/copyleft/fdl.html) tai CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], Wikimedia Commonsista.
Raunioiskasvit, Aspleniaceae: sukuja 23, lajeja n. 2780; kosmopoliittinen levinneisyys, puuttuu aavikoilta; Suomessa kampasaniainen (Blechnumspicant; harvinainen), muutama raunioislaji (Asplenium), kotkansiipi (Onoclea struthiopteris), kolme kiviyrttilajia (Woodsia), kolme metsäimarrelajia (Gymnocarpium), kaksi loikkolajia (Cystopteris), kaksi hiirenporraslajia (Athyrium), myyränporras (Diplazium sibiricum; harvinainen) ja kaksi nevaimarrelajia (Thelypteris). Hirvenkieli (Aspleniumscolopendrium). – Tekijä: Abalg [GFDL (http://www.gnu.org/copyleft/fdl.html) tai CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], lähde: Wikimedia Commons.
Kallioimarrekasvit, Polypodiaceae: n. 76 sukua, n. 4070 lajia; levinneisyys yleismaailmallinen; Suomessa kallioimarre (Polypodium vulgare), harvinaisena kaksi härkylälajia (Polystichum) sekä kuusi alvejuurilajia (Dryopteris). Kallioimarteen lehtiä. – Tekijä: H. Zell [GFDL (http://www.gnu.org/copyleft/fdl.html) tai CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], Wikimedia Commonsista.
Ketunlieon pystyhaaraista kasvustoa. – By Photo by Algirdas, Lithuania [Public domain], from Wikimedia CommonsNykyajan liekomaiset kasvit (Lycophyta) ovat pieni jäänne paljon loisteliaammasta menneisyydestä. Kivihiilikausi ajoittuu 359 – 299 miljoonan vuoden taakse noin 60 miljoonaa vuotta kestäneelle ajanjaksolle, ja siihen mennessä liekomaisten evoluutiolinja oli kehittynyt melkoisen monimuotoiseksi. Pienikokoisten ruohovartisten kasvien lisäksi niissä oli jättiläismäisiä puita, joiden rungon läpimitta saattoi paisua kahteen metriin ja korkeus nousta yli 40 metriin. Nämä jättiläiset kukoistivat miljoonia vuosia kosteikoissa, mutta kun ilmasto tuli kuivemmaksi permikaudelle (299 – 252 miljoonaa vuotta sitten) tultaessa, niiden diversiteetti väheni.
Pienikokoisia liekomaisia (eli lykofyyttejä) on säilynyt, ja nykyään niiden ryhmässä on noin 1200 tunnettua lajia, jotka muodostavat kolme heimoa: lieko-, mähkä- ja lahnanruohokasvit. Monet niistä kasvavat tropiikin puiden päällyskasveina käyttäen puita kasvualustanaan loisimatta niillä. Liekomaisia on myös lauhkean vyöhykkeen alueilla, ja useimmiten ne ovat metsien kenttäkerrosten kasveja, joskus jopa uposkasveja. Alkeisvarsikot voivat olla näkyvissä maan pinnalla ja ovat vihreitä ja yhteyttäviä, maansisäisäiset saavat ravintonsa sienirihmastoilta.
Riidenlieon perättömiä itiötähkiä. – Tekijä: Bernd Haynold [GFDL (http://www.gnu.org/copyleft/fdl.html), CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/) tai CC BY 2.5 (https://creativecommons.org/licenses/by/2.5)], Wikimedia CommonsistaVarsinainen liekokasvi edustaa itiösukupolvea eli on sporofyytti, jolla on enemmän tai vähemmän pystyt varret ja halkihaaraiset eli dikotomiset juuret. Lehdet ovat enimmäkseen pieniä, ja isommoimmissakin on vain yksi johtojänne (mikrofyllit). Mähkät kasvavat tavallisesti melko pienikokoisina vaakasuuntaisesti. Monilla niistä ja kaikilla lieoilla itiölehdet eli sporofyllit sijaitsevat nuijamaisina ja käpymäisinä rykelminä eli strobiluksina. Lahnanruohojen ainoa suku kasvaa soisilla kasvupaikoilla tai vedessä uposkasveina. Kaikkia näitä liekomaisten tyyppejä tavataan myös Suomessa.
Liekokasvien heimon (Lycopodiaceae) lajit ovat homosporisia, so. ne tuottavat vain yhdenlaisia itiöitä, joista itää kaksineuvoisia alkeisvarsikoita muna- ja siittiöpesäkkeineen (yksikotisuus). Sen sijaan mähkät (heimo Selaginellaceae) ja lahnanruohot (heimo Isoëtaceae) ovat heterosporisia: isoitiöistä kasvaa munapesäkkeellisiä alkeisvarsikoita ja pikkuitiöistä siittöpesäkkeellisiä; alkeisvarsikot eli gametofyytit ovat siten yksineuvoisia ja kaksikotisia.
Liekokasvien itiöt vapautuvat suurina määrinä yhdellä kertaa. Ne ovat hyvin öljypitoisia ja helposti syttyviä, joten mm. taikurit ja valokuvaajat ovat aikoinaan käyttäneet niitä savun ja valonvälähdysten aikaansaamiseen (ns. kärpäsruuti).
Suomessa kasvaa kuusi liekokasvien heimoon kuuluvaa lajia. Yleisimmät niistä ovat koko maassa kasvavat ketunlieko (Huperzia selago), riidenlieko (Lycopodium annotinum) ja keltalieko (Diphasiastrum complanatum). Ketunlieko voi kasvaa 20 cm pitkäksi ja on pystykasvuinen, ja sen itiöpesäkelehdet ovat kasvulehtien kaltaisia. Riidenlieon varsi voi olla yli metrin mittainen ja on suikertava ja liereä, mutta haarat ovat pystyjä, haarattomia tai niukkahaaraisia; itiötähkät sijaitsevat yksittäin ja ovat perättömiä. Keltalieon samanmittaset varret ovat litteitä, ja itiötähkät ovat perällisiä.
Keltalieko haaraisine itiötähkäperineen. – Tekijä: per.aasen (https://www.flickr.com/photos/candidum/3121381112/) [CC BY 2.0 (https://creativecommons.org/licenses/by/2.0)], lähde: Wikimedia CommonsMähkäkasvien heimosta maassamme esiintyy vain yksi laji, mähkä (Selaginella selaginoides). Se on yleinen vain Lapissa, eivätkä sen haarat ylitä 10 cm:ä. Vain itiöhaarat ovat pystyjä.
Mähkä. – Tekijä: Ivar Leidus [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], Wikimedia CommonsistaLahnanruohokasvien heimoon kuuluu maassamme kaksi yleisesti vesistöissä uposkasvina kasvavaa lajia: vaalea- (Isoëtes echinospora) ja tummalahnanruoho (I. lacustris). Niillä on pysty, lyhyt ja mukulamainen varsi, jossa kielekkeelliset lehdet ovat tiheänä ruusukkeena. Isoitiöpesäkkeet sijaitsevat uloimpien lehtien tyvituppien suojassa, pikkuitiöpesäkkeet keskimmäisten lehtien tyvellä.
Tummalahnanruoho. – Carl Lindman [Public domain tai Public domain], lähde: Wikimedia Commons
Olen kiipeilyssäni elämän puun oksistossa päässyt nyt Archaeplastida-haaran kohtaan, jossa kasveille on kehittynyt merkittävä innovaatio: kuljetussolukoita, joista syntyy johtojänteiksi nimitettyjä rakenteita. Näitä kasveja nimitetään putkilokasveiksi eli trakeofyyteiksi.
Muistutan mieleeni, että muut kolme aitotumaisten oksiston päähaaraa ovat nimeltään Excavata, SAR ja Unikonta. Ekskavaatteja ovat esim. silmäeliöt (Euglena). SAR-oksa (eli kreikkalaisperäisesti ’kladi’) saa nimensä kolmen haaransa etukirjaimista: Stramenopila (esim. ruskolevät), Alveolata (esim. ripsieliöt), Rhizaria (esim. huokoseliöt). Kolmas pääkladi on kiipeilyssäni vielä täysin koskemattomaksi jäänyt yksisiimaiset (Unikonta), joihin me ihmisetkin kuulumme yhtenä pienen pienenä oksanhaarana.
Oksisto haarautuu niin moneen kertaan yhä pienempiin osiin, että välillä tuntee olevansa eksyksissä ja suuret linjat hämärtyvät. Yritän kuitenkin pitää ne mielessäni ja hahmottaa niitä.
Yhteen ja samaan suureen oksaan kuuluvat liekomaiset, saniaiset ja siemenkasvit, ja kaikkien kolmen yhteisnimitys on putkilokasvit (Tracheophyta). Ne ovat kooltaan tavallisesti sammalia paljon isompia, minkä veden ja yhteyttämistuotteiden kuljetusjärjestelmä tekee mahdolliseksi. Mutta kuinka evoluutio ne synnytti?
Maakasvien eli alkiollisten kasvien sata miljoonaa ensimmäistä vuotta oli sammalten aikaa. Varhaisimmat putkilokasvifossiilit ajoittuvat 425 miljoonan vuoden taakse. Ne olivat itiökasveja, joilla oli kuitenkin hyvin kehittyneet kuljetussolukot. Näin saattoi kasvien kokokin kasvaa. Suvullinen lisäääntyminen oli vielä sammalten tasolla: siimalliset siittiöt uivat ympäristön vesikalvoissa munasoluja kohti, mikä vieläkin vangitsee lieot, kortteet ja saniaiset suhteellisen kosteisiin ympäristöihin.
Muinaiset putkilokasvien itiöpolvet eli sporofyytit olivat sammalista poiketen haaroittuneita ja riippumattomia sukusoluja tuottavista gametofyyteistä. Niiden korkeus jäi alle 20 senttimetrin. Haaroittumisen vuoksi ne pystyivät tuottamaan useampia itiöpesäkkeitä, sporangioita.
Esimerkkinä varhaisesta putkilokasvista mainittakoon Aglaophyton major. Sen fossiilit ovat 405 miljoonaa vuotta vanhoja, ja niiden mukaan se oli kasvutavaltaan halkihaarainen eli dikotominen. Haarojen päissä sijaitsivat itiöpesäkkeet. Sporofyytti oli siis monihaarainen toisin kuin sammalilla. Kasvilla oli sammalten tavoin juurtumahapsia, ritsoideja, ei oikeita juuria.
Aglaophyton major: itiöpesäke, itiöitä ja sporofyytti pesäkkeineen ja juurtumahapsineen. – By original: Falconaumanniderivative work: Peter coxhead [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)], via Wikimedia CommonsKun kasvit lisääntyivät tehokkaammin ja saattoivat kasvaa laajoiksikin kasvustoiksi, kilpailu valosta ja tilasta lisääntyi. Se oli omiaan lisäämään evoluution nopeutta, ja miljoonia vuosia kestänyt kehitys kohti metsäkasvillisuutta alkoi. Ensimmäiset metsät ilmaantuivat noin 385 miljoonaa vuotta sitten.
Varhaisilla putkilokasveilla oli joitakin samoja kehittyneitä ominaisuuksia kuin nykyisilläkin putkilokasveilla, mutta mm. juuria niillä ei ollut. Nykyisten putkilokasvien pääpiirteitä ovat sporofyytin hallitsevuus elinkierrossa, kuljetussolukot eli puu ja nila, hyvin kehittyneet juuret ja lehdet. Osa lehdistä on ns. sporofyllejä, joissa itiöpesäkkeet sijaitsevat.
Sporofyytti-vaiheen hallitsevuus elinkierrossa alkoi fossiileista päätellen kehittyä eräillä varhaisimmista putkilokasveista, joiden gameto- ja sporofyytit olivat kooltaan yhtäläisiä. Nykyisiin putkilokasviryhmiin edistyttäessä itiöpolvi on kehittynyt selvästi suuremmaksi ja rakenteeltaan monimutkaisemmaksi; pieniä sukusoluja muodostavia alkeisvarsikoita tuskin edes huomaa luonnossa; niitä on etsimällä etsittävä maasta tai jopa sen pinnan alta.
Voimakkaasti suurennettu lähikuva lehtivihreällisistä sanikkaisgametofyyteistä eli alkeisvarsikoista, joissa sukusolupesäkkeet sijaitsevat. – By Random Tree [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], from Wikimedia CommonsPutkilokasveilla on kahdenlaista kuljetussolukkoa: puuta eli ksyleemiä ja nilaa eli floeemia. Edellinen kuljettaa vettä ja mineraaleja juurista muihin kasvinosiin, jälkimmäinen taas vihreissä kasvinosissa syntyneitä yhteyttämistuotteita. Puusolukko koostuu putkimaisista putkisoluista eli trakeideista, joiden solunseinät ovat ligniini-nimisen polymeerin vahvistamia. Siitä johtuu puun kovuus, ja ligniini antaa kasville myös tukea, jotta se voi kasvaa kookkaaksikin, mistä on se etu, että pääsee paremmin valoon ja saa paremmin itiöt leviämään tuuleen.
Litulaukan (Alliaria petiolata) kuljetussolukkoa eli johtojänteitä poikittain leikattuna: puusolukko värjätty punaisella, nila sen ympärillä. – Tekijä: User:Micropix [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], Wikimedia CommonsistaJuurilla putkilokasvit imevät vettä ja ravinteita maaperästä ja ankkuroituvat siihen. Juuristo saattaa näin ollen kasvaa laajaksi. Nykyisten putkilokasvien juuren solukot muistuttavat varhaisten putkilokasvien varsien solukoita, mikä vihjaa siitä, että juuret ovat voineet kehittyä maata vasten olevista ja maan peittämiksi joutuneista varsista. Ilmeisesti juuria on kehittynyt monta eri kertaa eri putkilokasviryhmissä. Liekomaisten vanhimmilla fossiileilla oli jo noin 400 miljoonaa vuotta sitten yksinkertaisia juuria; saniaisten ja siemenkasvien kantamuodoilla sen sijaan ei niitä vielä ollut.
Lehdet huolehtivat pääosasta putkilokasvien fotosynteesissä eli hiilen yhteyttämisestä auringonvalon voimalla. Niitä on kahta tyyppiä: mikro- ja megafyllejä (suomeksi kirjaimellisesti pikku- ja isolehtiä). Edellisiä on vain liekomaisilla, ja niissä on vain yksi pituussuuntainen suoni, jonka muodostaa yksi johtojänne. Jäkimmäiset taas ovat suonitukseltaan monimutkaisempia ja hyvin haaroittuneita. Megafyllit voivat olla sekundaarisesti surkastuneita esim. monilla kuivien ympäristön kasveilla, jolloin varsi toimii yhteyttämiselimenä. Yleensä megafyllit ovat nimensä mukaan isompia ja tehokkaampia yhteyttämiselimiä kuin mikrofyllit. Mikrofyllejä ilmestyi fossiilistoon noin 410, megafyllejä noin 370 miljoonaa vuotta sitten devonikauden loppupuolella.
Itiöpesäkelehtien eli sporofyllien muodostuminen oli kasvien evoluutiossa merkittävä tapaus, sillä se itse asiassa mahdollisti kukkakasvienkin synnyn. Johtojänne- eli putkilokasveilla on rakenteeltaan vaihtelevia sporofyllejä: saniaisilla itiöpesäkkeet sijaitsevat tavallisesti pesäkelehden alapinnalla ryhmittyneinä ns. soruksiksi; liekomaisilla ja suurimmalla osalla paljassiemenisistä, esim. havupuilla, sporofyllit muodostavat käpymäisiä ryhmittymiä eli strobiluksia; kukkakasveilla eli koppisiemenisillä ne ovat hede- ja emilehtiä.
Itiötyyppien mukaan putkilokasvit ovat joko yhtäläis- tai erilaisitiöisiä (homo- tai heterosporisia). Suurin osa liekomaisista ja saniaisista on yhtäläisitiöisiä, so. niillä on vain yhdenlaisia itiöpesäkkeitä, jotka tuottavat yhdenlaisia itiöitä, joista kasvaa kaksineuvoisia (biseksuaalisia) alkeisvarsikoita muna- ja siittiöpesäkkeineen. Erilaisitiöisillä on puolestaan kahdenlaisia sporofyllejä, joista toiset kantavat isoitiöpesäkkeitä eli megasporangioita ja toiset pikkuitiöpesäkkeitä eli mikrosporangioita. Isoitiöistä kehittyy naarasalkeisvarsikoita ja pikkuitiöistä koirasalkeisvarsikoita.
Trakeofyytit eli putkilo- eli johtojännekasvit jaetaan luontevasti kolmeen ryhmään, joita voi pitää pääjaksoina (tieteellisen nimen pääte -phyta), luokkina (pääte -opsida) tai jonain muuna kategorioina luokittelijasta riippuen. Tässä ne ovat pääjaksomuodossa:
Liekomaiset, Lycophyta
Saniaiset, Monilophyta (sisältää myös kortteet, Equisetum)
Lehtisammalen elämänkierto eli sukupolven vuorottelu. Vasemmalta nuolien suuntaan: diploidi (kromosomisto kaksinkertaisena eli 2n) sporofyytti sijaitsee gametofyytin päässä, ja sen sporangiossa eli itiöpesäkkeessä tapahtuu meioosi eli haploideja itiöitä muodostava solunjako. Itiöt eli spoorit itävät ja kasvavat solunjakojen (mitoosien) seurauksena haploidiksi (kromosomisto yksinkertaisena) sukusoluja tuottavaksi gametofyytiksi. Koirasgametofyytti tuottaa siittiöpesäkkeessä eili anteridiossa siittiöitä, jotka kulkeutuvat vesipisaroiden mukana naaraskasvin munapesäkkeisiin eli arkegonioihin ja yhtyvät munasoluihin, jolloin tapahtuu hedelmöitys (fertilisaatio). Hedelmöitynyt munasolu jakautuu ja kasvaa alkioksi (embryo), josta sporofyytti kehittyy ja jää gametofyytin yhteyteen. – By Htpaul (I created this in Microsoft Paint) [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons
Lehtisammalet, Bryophyta
Lehtisammalten gametofyyttien eli sukusoluja tuottavien kasvien koko vaihtelee alle millimetristä 60 senttimetriin; yleensä ne eivät kuitenkaan ole 15 cm:ä pidempiä. Niiden lehtimäiset osat ovat tavallisesti vain yhden solukerroksen paksuisia, mutta karhunsammalilla (Polytrichum) ja niiden sukulaisilla voi niissä olla vahaisen pintakelmun eli kutikulan peittämiä harjanteita, mikä kertoo sopeutumisesta kuivahkoihin kasvupaikkoihin.
Lehtisammalten itiöitä tuottavat osat eli sporofyytit ovat tyypillisesti pitkäperäisiä ja paljain silmin havaittavia törröttäen vihreän gametofyytin jatkeena. Ne voivat olla gametofyyttejä pidempiä, jopa noin 20 cm pitkiä. Aluksi ne ovat lehtivihreällisiä ja yhteyttäviä, mutta kypsyessään muuttuvat ruskeiksi, jopa punertaviksi.
Lehtisammalia tunnetaan noin 15 000 lajia kaikkialta maapallolta, Suomesta on niitä löydetty vajaat 800. Lehtisammalten pääjakso Bryophyta voidaan ryhmitellä luokkiin ja lahkoihin seuraavasti (Ruggiero, M.A., Gordon, D.P., Orrell, T.M., Bailly, N., Bourgoin, T., Brusca, R.C., Cavalier-Smith, T., Guiry, M. D. & Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms. PLoS ONE 10(4): e0119248):
Luokassa Andreaeobryopsida on yksi lahko Andreaeobryales ja siinä vain yksi laji Andreaeobryum macrosporum Pohjois-Amerikan luoteisosissa.
Andreaeopsida-luokan muodostaa lahko Andreaeales yksin. Siihen kuuluu Suomessakin tavattava karstasammalten suku (Andreaea), joka on luokkansa ja lahkonsa ainoa.
Suurin lehisammalluokista on Bryopsida, jossa on 21 lahkoa. Suomen sammalista siihen kuuluvat mm. omenasammalet (Bartramia), hiirensammalet (Bryum), harmosammalet (Hedwigia), palmikkosammalet (Hypnum), hiippasammalet (Orthotrichum), sompasammalet (Splachnum), kaviosammalet (Buxbaumia), kuulasammal (Archidium alternifolium), kynsisammalet (Dicranum), kivisammalet (Grimmia), munasammal (Diphyscium foliosum), kellosammalet (Encalypta), nuotiosammal (Funaria hygrometrica), tuppisammalet (Timmia) ja kiirunansammal (Oedipodium griffithianum).
Luokkaan Polytrichopsida kuuluvat karhunsammalet (Polytrichum), joita maassamme on kymmenen lajia. Lahkoja luokassa on vain yksi.
Sphagnopsida-luokka käsittää kaksi lahkoa: Ambuchananiales ja Sphagnales. Jälkimmäiseen kuuluvat maamme 42 rahkasammallajia (Sphagnum), jotka ovat soidemme valtasammalia.
Luokan Takakiopsida muodostaa Takakiales-lahko yksinään, ja siinäkin on vain yksi 2-lajinen suku (Takakia), joka kasvaa Pohjois-Amerikan länsiosissa sekä Keski- ja Itä-Aasiassa.
Luokassa Tetraphidopsida on niin ikään yksi lahko: Tetraphidales. Sen ainoaan heimoon kuuluu kaksi sukua, joissa on yhteensä neljä lajia. Maassamme kasvaa lahosammal (Tetraphis pellucida).
Pahtaomenasammal (Bartramia halleriana). – By HermannSchachner [CC0], from Wikimedia Commons.Sirohiirensammal (Bryum elegans). – By HermannSchachner [CC0], from Wikimedia Commons.Kiviharmosammal (Hedwigia ciliata). – By Kurt Stüber [1] [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/)], via Wikimedia Commons.Kalliopalmikkosammal (Hypnum cupressiforme). – By Kurt Stueber (www.biolib.de) [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/)], via Wikimedia Commons.Isohiippasammal (Orthotrichum lyellii). – By HermannSchachner [CC0], from Wikimedia Commons.Jouhisompasammal (Splachnum sphaericum). – By HermannSchachner [CC0], from Wikimedia Commons.Korpikarhunsammalia (Polytrichum commune), oikealla itiöpesäkkeitä perineen eli sporofyyttejä. – By Kristian Peters — Fabelfroh 14:35, 21 October 2007 (UTC) (photographed by Kristian Peters) [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC-BY-SA-3.0 (http://creativecommons.org/licenses/by-sa/3.0/)], via Wikimedia Commons.Sararahkasammal (Sphagnum fallax) on maassamme yleinen, pohjoisessa harvinaisempi. – By Bernd Haynold [GFDL (http://www.gnu.org/copyleft/fdl.html) or CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], from Wikimedia Commons.Lahosammal (Tetraphis pellucida) edustaa luokkaa Tetraphidopsida. – James Lindsey at Ecology of Commanster [CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5) or CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons.
Sarvisammalet
Sarvisammalet on sammalpääjaksoista pienin sisältäen vain 100 lajia. Ne ovat sammalista putkilokasvien (liekomaiset, saniaiset, siemenkasvit) lähimpiä sukulaisia.
Sekä suomalainen että pääjakson tieteellinen nimi Anthocerophyta viittaavat sporofyytin sarvimaiseen muotoon. Se voi kasvaa viiden cm:n mittaiseksi. Toisin kuin maksa- ja lehtisammalilla siinä ei ole vartta eli perää, vaan se on kokonaan itiöpesäkettä. Itiöt vapautuvat, kun pesäke halkeaa ylhäältä alkaen.
Gametofyytit eli suvullisesti lisääntyvä sukupolvi on tavallisesti vain sentin tai kahden levyinen ja kasvaa vaakatasossa. Yhdessä gametofyytissä voi olla kiinnittyneenä useita itiöpesäkesarvia.
Sarvisammalet ovat usein avoimien ja kosteiden paikkojen pioneerikasveja. Kyseiset kasvupaikat ovat vähätyppisiä, mutta sarvisammalilla on symbiontteinaan typpeä sitovia syanobakteereja eli ns. sinileviä.
Sarvisammalet jaetaan kahteen luokkaan: Anthocerotopsida ja Leiosporocerotopsida. Edellisessä on neljä lahkoa, jälkimmäisessä vai yksi. Lahkot ovat Anthocerotales, Dendrocerotales, Phymatocerotales, Notothyladales ja Leiosporocerotales (Ruggiero, M.A., Gordon, D.P., Orrell, T.M., Bailly, N., Bourgoin, T., Brusca, R.C., Cavalier-Smith, T., Guiry, M. D. & Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms. PLoS ONE 10(4): e0119248.)
Sarvisammal (Anthoceros agrestis) on toinen maassamme tavattavista kahdesta sarvisammallajista (toinen on puikkosammal, Phaeoceros carolinianus). – By BerndH [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0) or GFDL (http://www.gnu.org/copyleft/fdl.html)], from Wikimedia Commons.
.
Fylogeneettiset eli polveutumista koskevat tutkimukset osoittavat että kaikki kolme sammalten pääryhmää, maksa-, lehti- ja sarvisammalet, erilaistuivat omiksi kladeikseen maakasvien evoluution varhaisvaiheessa. Nämä kladit seurasivat toisiaan siten, että maksasammalet erilaistuivat ensin ja lehti- ja sarvisammalet myöhemmin yhteisestä kantamuodosta, joka oli myös putkilokasvien kantamuoto. Sarvisammalilla ja putkilokasveilla on ollut nuorempi kantamuoto kuin lehti- ja sarvisammalilla.
Fossiilistosta on löydettävissä tukea tälle ajatukselle. Varhaisimmat kasvien itiöt 470-450 miljoonan vuoden takaa ovat rakenteeltaan nykyisten maksasammalten itiöiden kaltaisia. Noin 430 miljoonaa vuotta vanhoihin kerrostumiin ilmestyy myös lehti- ja sarvisammalten itiöitä muistuttavia itiöitä. Melko pian tämän jälkeen ilmestyivät jo ensimmäiset putkilokasvit 425 miljoonaa vuotta sitten.
Sammalien elinkierron hallitseva vaihe on kromosomistoltaan yksiannoksinen eli haploidi gametofyytti eli suvullinen sukupolvi. Se on isompi ja pitkäikäisempi kuin sporofyytti eli suvuton sukupolvi, joka on diploidi, sillä se syntyy sukusolujen yhtymisen seurauksena. Sporofyytin vapauttamat itiöt itävät sopivissa olosuhteissa ja kasvavat gametofyytiksi. Itiöt muodostuvat meioottisesti, so. niitä muodostavissa solujaoissa tapahtuu vähennysjako, jolloin kromosomiston eri puolikkaat joutuvat eri itiöihin, meiosporeihin. Itävä itiö muodostaa aluksi alkeisrihman, protoneeman, joka on lehtivihreällinen, haarainen ja yhden solukerroksen paksuinen. Sen laaja pinta absorboi vettä ja mineraaleja, ja suotuisissa olosuhteissa se tuottaa silmumaisia muodostumia. Kussakin niistä on kärkikasvusolukko (apikaalimeristeemi), josta syntyy sukusoluja tuottava gametoforiksi nimitetty rakenne. Protoneema ja yksi tai useampia gametoforeja kehittyy sammalen gametofyytiksi, versoksi, jota yleisesti pidämme varsinaisena sammalkasvina. – Kuvia.
Sammalet ovat pienikokoisia, joskus jopa vähäpätöisiä. Niillä ei ole tukirakenteita eikä kuljetussolukkoja eli johtojänteitä, jotka voisivat kuljettaa vettä ja ravinteita kasvin eri osiin, joten ne eivät voi kasvaa korkeiksi. Joillakin sammalilla on kuljetussolukkomaisia rakenteita varren keskiosassa, mikä sallii kasvin kasvavan jopa 60 cm korkeaksi. Nämä rakenteet ovat syntyneet riippumatta putkilokasveille kehittyneistä johtojänteistä aivan itsenäisesti ja ovat esimerkki rinnakkaisevoluutiosta eli konvergenssista.
Sammalkasvi kiinnittyy alustaansa juurtumahapsilla eli ritsoideilla, jotka ovat maksa- ja sarvisammalilla yksisoluisia, lehtisammalilla monisoluisia rihmoja. Niissä ei siten ole putkilokasvien juurille ominaisia erilaistuneita solukoita. Ritsoidit ovat pelkästää kiinnittymiselimiä eivätkä toimi ensisijaisina veden ja mineraalien kerääjinä.
Gametofyytti synnyttää suojaavan solukon alle gametangioita eli sukusolupesäkkeitä. Pullomainen arkegonio eli munapesäke sisältää yhden munasolun, joka sijaitseen pullon pohjalla, jonne pitkänomainen kaula johtaa. Anteridio eli siittiöpesäke tuottaa sitä vastoin joukon siittiöitä. Joskus arkegonio ja anteridio sijaitsevat samassa sammalyksilössä, jolloin sammal on kaksineuvoinen, mutta yleisemmin sammalgametofyytissä on vain jompia kumpia sukusolupesäkkeitä, jolloin ne ovat yksineuvoisia, koiraita tai naaraita. Jotta siittiöt voivat saavuttaa munasolun, tarvitaan vettä, jonka kalvossa siimalliset siittiöt uivat arkegoniota kohti sen erittämien aineiden houkuttamina. Munasolu pysyy aloillaan arkegonion pohjalla, ja sinne ehtinyt siittiö hedelmöittää sen. Hedelmöittynyt munasolu eli tsygootti jää paikalleen arkegonioon, josta muodostuu alkiota ravitseva solukko. Sen turvin alkio kehittyy sporofyytiksi.
Alkiosta kehittyvä sporofyytti on aluksi vihreä ja yhteyttävä, vaikkei eroakaan gametofyytistä eikä elä siitä riippumatta. Sammalsporofyytti jää koko iäkseen gametofyytin yhteyteen ja on siitä riippuvainen.
Suvullisen lisääntymisen lisäksi useilla sammalilla on kasvullista, suvutonta lisääntymistä, jossa menetelmät vaihtelevat. Sammal saattaa esimerkiksi kasvattaa itusilmumaisia sivuhaaroja, jotka irtoavat emokasvista ja muodostavat kloonin kyseiselle kasvupaikalle. Kloonissa kaikki yksilöt ovat geneettisesti identtisiä.
maksasammalille ominaista
Maksasammalten suomalainen ja tieteellinen nimi Hepatophyta (kreikan hepar, maksa) johtuu joidenkin lajien maksan liuskaisuutta muistuttavasta gametofyytistä (esim. keuhkosammalet, Marchantia; kuvia). Keskiaikaisen ns. signatuuriopin mukaan ajateltiin kyseisten maksasammalten sopivan maksasairauksien hoitoon. Signatuuriopin mukaan kunkin elimen sairauksiin sopi lääkkeeksi kasvi, joka tai jonka jokin osa on kyseisen elimen muotoinen.
Monien maksasammalten – esim. keuhkosammalen – ulkonäkö tuo mieleen monisoluisen viherlevän litteän ja sekovarsimaisen gametofyytin vuoksi. Keuhkosammalilla sekovarresta nousee palmumaisia gametangioita eli sukusolupesäkkeitä. Suurennuslasin avulla voi nähdä varrelliset sporofyytit, jotka kehittyvät sukusolupesäkkeen liuskojen alapinnalle. Kypsyneet itiöt vapautuvat varren päässä olevasta pyöreästä itiöpesäkkeestä.
Maksasammaliin kuuluu myös enemmän lehtisammalia muistuttavia lajeja, joilla on lehtimäisiä ulokkeita kiinnittyneenä kasvin varsimaiseen osaan (esim. kastesammalet, Plagiochila; kuvia). Tämäntyyppisiä on maksasammalten enemmistö. Niiden lehdet ovat kiinnittyneet kolmeen riviin. Useimmiten verson alapinnan lehdet ovat muita pienempiä, ja niitä kutsutaan vatsalehdiksi; varren sivulla ovat isommat kylkilehdet.
maksasammalten luokittelu
Maksasammalten noin 9000 lajia ryhmitellään nykyään kolmeen luokkaan: Haplomitriopsida, Jungermanniopsida (enemmistö maksasammalista) ja Marchantiopsida. Kasvien luokittelussa (systematiikassa ja taksonomiassa) käytetään kansainvälisen nimistösäännostön mukaisia kategorioita, jotka tunnistaa tieteellisen nimen loppuosasta. Pääjakson (eli kaaren, tai sitä ylemmän taksonin) pääte on –phyta, luokan –opsida, alaluokan –idae, lahkon –ales ja heimon –aceae. Kantasanaksi otetaan ns. nimisuku, esim. Marchantia (keuhkosammalet), jonka taivutusvartaloon Marchanti– lisätään em. päätteitä sen mukaan, puhutaanko suvun sisältämästä luokasta, lahkosta tai heimosta tai muusta taksonista.
Systematiikassa pohditaan eliöiden sukulaisuussuhteista, ja taksonomia tarkoittaa niiden mukaisten ryhmien nimeämistä. Usein kuitenkin fylogeneettisiä sukuhaaroja nimetään pohtimatta varsinaista systemaattista kategoriaa, jolloin nimetty evoluutiopuun haara ei noudata em. kategorian osoittavaa sääntöä. Esimerkiksi kasvit kuuluvat yhdessä punalevien kanssa kehityshaaraan, joka kantaa nimeä Archaeplastida, ja siihen kuuluvaan alahaaraan Viridiplantae ja edelleen siihen kuuluvaan oksaan Streptophyta (nimi vaikuttaa pääjaksolta!). Viimeksi mainittu evoluutiopuun oksa haarautuu eräisiin ennen viherleviin luettuihin haaroihin (mm. näkinpartaiset, Charophyta) ja alkiollisten kasvien eli versokasvien haaraan Embryophyta, jota voi pitää kasvikuntana suppeassa merkityksessä (laajimmassa merkityksessä kasvikunta olisi koko Archaeplastida-ryhmä).
Seuraavassa taulukossa on lueteltu maksasammalten luokat, alaluokat, lahkot ja heimot, mutta alalahkot on jätetty huomiotta (Söderström ja muut 2016).
Kaari (pääjakso) Hepatophyta eli Marchantiophyta, maksasammalet
Luokka Haplomitriopsida
Alaluokka Haplomitriidae
Lahko Calobryales. Yksi heimo: Haplomitriaceae. – kuvia
Alaluokka Treubiidae
Lahko Treubiales. Yksi heimo: Treubiaceae. – kuvia
Lahko Porellales. Seitsemän heimoa: Frullaniaceae, Jubulaceae, Lejeuneaceae, Goebeliellaceae, Lepidolaenaceae, Porellaceae, Radulaceae. – kuvia
Lahko Ptilidiales. Kolme heimoa: Herzogianthaceae, Neotrichocoleaceae, Ptilidiaceae. – kuvia
Alaluokka Metzgeriidae
Lahko Metzgeriales. Kaksi heimoa: Aneuraceae, Metzgeriaceae. – kuvia
Lahko Pleuroziales. Yksi heimo: Pleuroziaceae, jossa suku Pleurozia(kuvia), jota ei tule sekoittaa lehtisammaliin kuuluviin seinäsammaliin (Pleurozium)!
Alaluokka Pelliidae
Lahko Fossombroniales. Viisi heimoa: Allisoniaceae, Calyculariaceae, Fossombroniaceae, Makinoaceae, Petalophyllaceae. – kuvia
Lahko Pallaviciniales. Viisi heimoa: Hymenophytaceae, Moerckiaceae, Pallaviciniaceae, Phyllothalliaceae, Sandeothallaceae. – kuvia
Lahko Pelliales. Kaksi heimoa: Noterocladaceae, Pelliaceae. – kuvia
Luokka Marchantiopsida
Alaluokka Blasiidae
Lahko Blasiales. Kaksi heimoa, joista jälkimmäinen on hävinnyt: Blasiaceae, †Treubiitaceae. Blasiaceae-sammalissa on symbiontteina Nostoc-syanobakteereita (ns. sinileviä). – kuvia
Alaluokka Marchantiidae
Lahko Lunulariales. Yksi heimo: Lunulariaceae. – kuvia
Lahko Marchantiales. 13 heimoa: Aytoniaceae, Cleveaceae, Conocephalaceae, Corsiniaceae, Cyathodiaceae, Dumortieraceae, Marchantiaceae, Monocleaceae, Monosoleniaceae, Oxymitraceae, Ricciaceae, Targioniaceae, Wiesnerellaceae. – kuvia
Lahko Neohodgsoniales. Yksi heimo: Neohodgsoniaceae. – kuvia
Lahko Sphaerocarpales. Neljä heimoa, joista yksi on hävinnyt: Monocarpaceae, †Naiaditaceae, Riellaceae, Sphaerocarpaceae. – kuvia
Lähde: Söderström, Lars, Anders Hagborg, Matt von Konrat, Sharon Bartholomew-Began, David Bell, Laura Briscoe, Elizabeth Brown, D. Christine Cargill, Denise P. Costa, Barbara J. Crandall-Stotler, Endymion D. Cooper, Gregorio Dauphin, John J. Engel, Kathrin Feldberg, David Glenny, S. Robbert Gradstein, Xiaolan He, Jochen Heinrichs, Jörn Hentschel, Anna Luiza Ilkiu-Borges, Tomoyuki Katagiri, Nadezhda A. Konstantinova, Juan Larraín, David G. Long, Martin Nebel, Tamás Pócs, Felisa Puche, Elena Reiner-Drehwald, Matt A. M. Renner, Andrea Sass-Gyarmati, Alfons Schäfer-Verwimp, José Gabriel Segarra Moragues, Raymond E. Stotler, Phiangphak Sukkharak, Barbara M. Thiers, Jaime Uribe, Jiří Váňa, Juan Carlos Villarreal, Martin Wigginton, Li Zhang, Rui-Liang Zhu. (2016) World checklist of hornwortsand liverwortsPhytoKeys 59:1–828. Full textReference page.
Haplomitriopsida-luokka käsittää vain suvut Treubia(kuvia), Apotreubia(kuvia) ja Haplomitrium(kuvia), joissa on yhteensä 15 lajia, jotka kasvavat pääasiassa Australaasiassa. Viimeaikaiset tuman, mitokondrioiden ja plastidien DNA:n kladistiset analyysit sijoittavat tämän monofyleettisen ryhmän kaikkien muiden maksasammalten sisarryhmäksi maksasammalten evoluutiopuussa. Ryhmä tarjoaa sen vuoksi näkymän sekä maksasammalten että alkiollisten kasvien varhaiseen evoluutioon. Treubia-suvun kuusi lajia kasvaa alustanmyötäisinä lehdellisinä sekovarsina; lehdet voivat olla siipimäisesti tai laskostuneet ylöspäin yhteen. Suvussa on piirteitä, jotka eroavat muista sammalista; poikkeuksellista on mm. varren keskisuonen rakenne. Haplomitrium-lajeilla on maanalainen varsi, josta kohoaa näkyviin pyöreälehtisiä haaroja. Muista maksasammalista poiketen tämän suvun kasveilla ei ole juurtumahapsia, ja varsissa on keskellä vettä kuljettava rakenne. Sen ympärillä on lieriö, joka kuljettaa ravinteita. Nämä rakenteet tuovat mieleen putkilokasvien puu- ja nilasolukot.
Luokka Jungermanniopsida on suurin maksasammalten kolmesta luokasta. Siihen kuuluvat edellä kuvatut lehtisammalmaiset muodot, joilla on kolme riviä lehtimäisiä ulokkeita: kaksi ns. kylkilehtiriviä ja alapinnan vatsalehtirivi.
Marchantiopsida-luokan lajien sekovarsi on levymäinen, usein halkihaarainen ja leveäliuskainen. Solukkorakenne on pitkälle erikoistunut, ja niillä on solurykelmiä, joiden soluissa on kalvon ympäröimiä oljyrakkuloita.
Cooksonia oli yksi varhaisimmista alkiollisista kasveista. Kuppimaiset varrenkärjet ovat itiöpesäkkeitä. – MUSE [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)].Läheisimmät versokasvien leväsukulaiset ovat paljolti yksisoluisia ja solukolonioita muodostavia. On siten todennäköistä, että ensimmäiset versokasvitkin olivat pieniä, ja siksi varhaisimpien maakasvien fossiilien etsintä on keskittynyt mikrofossiileihin. Niitä varhaisemmat mikroskooppiset eliöt alkoivat maanvalloituksen jo 1 200 miljoonaa vuotta sitten. Noin 470 miljoonaa vuotta sitten niiden joukkoon ilmestyi uutta: varhaisten alkiollisten kasvien itiöitä, jotka eroavat levien ja sienten itiöistä.
Yksi erottava tekijä levien ja sienten itiöihin nähden on kasvi-itiöiden kemiallinen koostumus. Se näet vastaa nykyisten itiökasvien itiöiden koostumusta ja on erilainen, kuin muilla eliöillä. Toisekseen muinaisten kasvien itiöiden seinämät ovat rakenteeltaan samanlaisia kuin nykyisillä maksasammalilla (Marchantiophyta). 450 miljoonan vuoden ikäisistä kalliokerrostumista on löytynyt itiöitä kasvien pintarakenteeseen, kutikulaan, uponneina, ja se muistuttaa nykyisten kasvien itiöpesäkkeiden solukkoa. 425 miljoonan vuoden takaa tunnetaan isompien kasvirakenteiden fosiilleja, mm. yläkuvassa olevan Cooksonian sporangio eli itiöpesäke.
Nykyisten alkiollisten kasvien (Embryophyta) eri ryhmien evoluutio ja nykyään tunnetujen lajien määrä:
Noin 470 milj. v. sitten alkiollisten eli versokasvien synty: putkilottomat kasvit, joita nykyään edustavat sammalet
Maksasammalet (Marchantiophyta eli Hepatophyta), 9000 lajia – kuvia
Gnetales (luumio-, efedra- ja ikilehtikasvit), 75 lajia – kuvia
Koppisiemeniset (Angiospermae), 250 000 lajia – kuvia
Kasvit voidaan jaotella rakennetasojen mukaan mm. sen perusteella, onko niillä kuljetussolukkoa vai ei. Kuljetussolukko on putkeksi yhtyneitten solujen (ja niihin mahdollisesti liittyvien muiden solujen) muodostama veden ja ravinteiden kuljetusjärjestelmä. Useimmilla nykykasveilla se on, ja niitä nimitetään putkilokasveiksi veden kuljetukseen sopeutuneitten putkimaisten rakenteiden mukaan. Sammalilla ei ole varsinaisia putkiloita, vaikkakin joillakin niistä on yksinkertainen kuljetussolukko. Sammalet eli bryofyytit eivät kuitenkaan ole monofyleettinen kladi eli kehityshaara, minkä molekyylitutkimukset ja siittiön morfologia osoittavat. Kolme eri sammalryhmää seuraavat evoluutiopuussa toisiaan erillisinä kladeina eli ne muodostavat parafyleettisen ryhmän.
Putkilokasvit muodostavat sitä vastoin monofyleettisen kladin, jossa on mukana niiden kantamuoto ja kaikki sen jälkeläiset. Valtaosa nykyisistä kasveista, yli 90%, kuuluu tähän kladiin. Kladin ensimmäiset haarat ovat liekomaiset ja niitä seuraavat saniaismaiset, joille on yhteistä se, että niiltä puuttuvat siemenet; ne ovat siis itiökasveja. Nekin muodostavat sammalten tavoin rakennetason (gradus), eivät kladia, sillä saniaismaisilla ja siemenkasveilla on nuorempi yhteinen kantamuoto kuin liekomaisilla ja saniaismaisilla.
Siemenkasvien kladi käsittää nykyisistä kasveista valtaosan. Siemen on alkio, joka on saanut suojakseen kuoren ja mukaansa ravinnesolukon, jonka varassa alkio kehittyy taimeksi. Siemenkasvit jakautuvat kahteen kladiin ajatellen nykyään elossa olevia lajeja. Nykyään elävillä paljassiemenisillä (mm. havupuut) siemen on paljaana emilehden pinnalla, kun taas kukkakasveilla eli koppisiemenisillä se on emilehden muodostaman kammion sisällä ikään kuin kopissa ja siten paremmassa suojassa. Lähes 90% nykyisistä kasvilajeista on koppisiemenisiä.
On vielä huomattava, että näiden nykyään elävien kasviryhmien lisäksi tunnetaan joukko fossiilisia sukupuuttoon kuolleita ryhmiä, joita ei edellisessä kuvauksessa ole otettu huomioon. Nämä fossiilit paljastavat välimuotosarjoja niiden kasviryhmien väliin, joita on nykyisellä maaplaneetalla.
Maksasammalet ovat alkukantaisin ryhmä nykyisistä alkiollisista kasveista (Embryophyta). – Ernst Haeckel [Public domain].Maapallomme ei aina ole ollut vihreä. Kesti miljardeja vuosia planeettamme synnystä ennenkuin ensimmäinen viherrys ilmaantui kuivalle maalle syanobakteerien ja aitotumaisten alkueliöiden muodostamana ohuena ja kalvomaisena peitteenä noin 1,2 miljardia vuotta sitten. Vasta 500 miljoonaa vuotta sitten pienet kasvit yhdessä sienten ja eläinten kanssa liittyivät niiden seuraan. Ensimmäiset metsät ilmestyivät noin 385 miljoonaa vuotta sitten.
Versokasveja eli alkiollisia kasveja (Embryophyta), juuri niitä, joita varsinaisesti miellämme kasveiksi, on nykyään lähes 300 tuhatta lajia. Ne ovat sopeutuneet lähes kaikkiin olosuhteisiin maapallolla, ja osa on palannut takaisin vesielämään. Versokasvit mahdollistivat muiden elämänmuotojen maanvalloituksen lisäämällä ilmakehän happipitoisuutta ja muodostamalla maaeläimille ravinnonlähteen. Kasvit myös ovat muodostaneet erilaisia elinympäristöjä, kuten aroja ja metsiä, ja meidän ihmisten mielessä ne enimmäkseen määräävät maiseman.
Maanvalloitus vaati kasveilta evolutiivisia innovaatioita
Versokasvien kantamuodot eli eräät karofyytteihin kuuluvat viherlevät kasvoivat järvien ja lampien matalissa rantavesissä ja joutuivat kestämään silloin tällöin kuivuutta. Niissä oloissa luonnonvalinta suosii leväyksilöitä, jotka kestävät parhaiten kuivumista. Sporopolleniini on kestävä polymeeri, ja sen muodostama kerros estää tehokkaasti kuivumista. Karofyyttien tsygootit eli hedelmöityneet munasolut saivat sellaisen peitteen, ja sama aine suojaa maakasvien itiöitä.
Riippumattomuus vedenalaisuudesta avasi uusia mahdollisuuksia ensimmäisille maakasveille. Auringonvaloa sai nyt runsaammin, kun vesi ei ollut sitä taittamassa eikä plankton varjostamassa, ilmakehässä on runsaammin hiilidioksidia kuin vedessä, ja kosteat rannat olivat ravinteikkaita. Yhteyttäminen tehostui.
Jotta näistä eduista pääsi nauttimaan, oli kohdattava muutamia haasteita. Veden vähyys ja sen suoma tuki ja noste poistuivat. Oli kohdattava kuivuus ja maan vetovoima eli tarvittiin vedenhankinta ja kuljetusjärjestelmä sekä tukirakenteita, jotta voitiin kasvaa maasta ylöspäin. Kasvit erilaistuivat ja monipuolistuivat, kun syntyi uusia sopeutumia eli adaptaatioita, jotka mahdollistivat menestymisen vaikeuksista huolimatta.
Versokasvien systemaattinen asema
Bakteerit, Bacteria
Arkeonit, Archaea
Aitotumaiset, Eukarya
Ekskavaatit, Excavata (mm. silmälevät)
SAR (mm. ruskolevät)
Archaeplastida
Punalevät, Rhodophyta
Glaukofyytit, Glaucophyta
Viridiplantae
Viherlevät, Chlorophyta
Streptofyytit, Streptophyta
Kuusi karofyyttiryhmää, mm. näkinpartaiset (Charophyceae)
Versokasvit, Embryophyta
Yksisiimaiset, Unikonta (mm. sienet ja eläimet)
Versokasvit
Kun verso- eli maa- eli alkiolliset kasvit (Embryophyta; kreikan embryo ’alkio’ ja fyton ’kasvi’) kehittyivät, syntyi uusia adaptaatioita kuivalla maalla selviytymiseksi ja lisääntymiseksi. Kantamuodolla ei ollut mm. sukupolvenvuorottelua, monisoluista alkiota, itiöpesäkkeiden eli sporangioiden tuottamia kestäviä itiöitä, monisoluisia gametangioita eli sukusolupesäkkeitä eikä kärkikasvusolukkoa (apikaalimeristeemiä). Nämä ominaisuudet kehittyivät vasta versokasveille, vaikka kaikilla niiden edustajilla ei ole näitä ominaisuuksia: joiltakin kasveilta jotkin ominaiset ovat sekundaarisesti kadonneet.
sukupolvenvuorottelu
Versokasveilla on kaksi sukupolvea elinkierrossaan: sukusoluja tuottava gametofyytti ja suvuttomia itiöitä tuottava sporofyytti. Vastaavanlainen sukupolvenvuorottelu oli jo kehittynyt monissa leväryhmissä, mutta ei karofyyteillä, joihin versokasvien kantamuoto kuului.
Kasvien sukupolvenvuorottelua ei tule sekoittaa muiden suvullisesti lisääntyvien eliöiden elinkierrossa oleviin haploidiin ja diploidiin vaiheeseen. Edellisessä soluissa on vain yksi kromosomiannos (merkitään kirjaimella n), jälkimmäisessä kaksi kromosomiannosta (2n). Esim. ihmisellä ja muilla eläimillä vain sukusolut ovat haploideja, kaikki muu tsygootista alkaen diploidia elämänvaihetta. Kasveilla sen sijaan esiintyy sekä monisoluinen haploidi organismi että monisoluinen diploidi organismi. Edellinen on gametofyytti, joka tuottaa siittiöitä ja/tai munasoluja, jotka yhtyessään tuottavat diploideja tsygootteja. Tsygootti alkaa jakautua mitoottisesti, mikä tarkoittaa, että tytärsolut säilyvät diploideina, ja lopulta muodostuu diploidi monisoluinen sporofyytti (2n). Sporofyytti tuottaa meioottisen solujaon eli vähennysjaon välityksellä haploideja itiöitä (haplosporeja, n). Itiöt itävät ja kasvavat gametofyyteiksi (n), jotka ovat yleensä huomiota herättämättömämpiä kuin sporofyytit. Gametofyytit voivat olla koiraita tai naaraita tai kaksineuvoisia.
alkio (embryo)
Hedelmöitynyt munasolu eli tsygootti kehittyy monisoluiseksi alkioksi, joka jää osaksi naarasgametofyyttiä ja siitä riippuvaiseksi. Emokasvin solukot suojelevat kehittyvää alkiota ja ravitsevat sitä mm. sokereilla ja aminohapoilla. Alkiossa on erikoistuneita soluja, joiden kautta ravinteet välittyvät emokasvista. Tällainen monisoluinen ja emokasvista riippuva alkio on tyypillinen vain embryofyyteille.
Itiöt
Kasvien itiöt ovat haploideja (n) lisääntymissoluja, jotka voivat kasvaa monisoluisiksi haploideiksi gametofyyteiksi mitoottisesti. Polymeeri nimeltään sporopolleniini tekee itiöiden seinämät koviksi ja ympäristön paineita kestäviksi. Tämä kemiallinen sopeuma tekee itiöt kykeneviksi leviämään kuivassa ilmassa niiden vahingoittumatta.
Sporofyytissä on monisoluisia itiöpesäkkeitä eli sporangioita, jotka tuottavat itiöitä (spooreja). Sporangiossa diploidit (2n) sporosyytit eli itiöiden emosolut käyvät läpi vähennysjaon (meioosin) ja tuottavat siten haploideja itiöitä. Sporangion ulommat solukot suojaavat kehittyviä itiöitä, kunnes ne vapautuvat ilmaan. Monisoluiset itiöpesäkkeet, jotka tuottavat sporopolleniiniseinäisiä itiöitä, ovat kasvien avainsopeutuma maaelämään. Vaikka karofyytitkin tuottavat itiöitä, ei niillä ole monisoluisia pesäkkeitä eikä niiden siimallisissa ja vedessä leviävissä itiöissä ole sporopolleniinia.
monisoluiset gametangiot
Maakasvit tuottavat sukusolunsa monisoluisissa sukusolupesäkkeissä eli gametangioissa. Munapesäkkeitä nimitetään arkegonioiksi, ja ne ovat päärynänmuotoisia, ja niissä syntyy yksi liikkumaton munasolu, joka jää arkegonion sisään. Siittiöpesäkkeet eli anteridiot tuottavat siittiöitä, jotka vapautuvat ympäristöönsä. Monissa kasviryhmissä siittiöt ovat siimallisia ja uivat arkegoniota ja munasolua kohti vesipisaroiden tai vesikalvon välityksellä. Hedelmöitys tapahtuu arkegoniossa, jossa tsygootti kehittyy alkioksi. Siemenkasveilla gametofyytit ovat niin surkastuneet kooltaan, että arkegoniot ja anteridiot puuttuvat monista niiden kehityslinjoista.
kärkikasvusolukko
Kuivalla maalla fotosynteettinen eliö saa tärkeät resurssinsa kahdesta aivan erilaisesta lähteestä. Valo ja hiilidioksidi saadaan pääasiassa ilmakehästä, vesi ja mineraalit puolestaan maaperästä. Useimmilla kasveilla on juuret ja ilmaversot, jotka voivat kasvaa pituutta lisäten siten mahdollisuuksia saavuttaa tarvittavia voimavaroja. Pituuskasvusta huolehtivat koko kasvin elinajan kärkikasvusolukot eli apikaalimeristeemit, jotka ovat verson kärkien kasvupisteitä: niissä yksi tai useampi solu jakautuu jatkuvasti tai ainakin toistuvasti. Tuotetut uudet solut erilaistuvat pinta- eli epidermisoluiksi, jotka muodostavat kasvin ulkopinnan, sekä muiksi, verson sisäisiksi solukoiksi. Ilmaverson apikaalimeristeemi tuottaa myös lehdet useimmilla kasveilla.
muita adaptaatioita
Em. viiden tärkeimmän sopeutuman lisäksi kasveilla on muitakin kehittyneitä ominaisuuksia, jotka ovat adaptaatioita elämään kuivalla maalla. Monilla on esim. pintasolukon eli epidermin päällä peite, kutikula, joka muodostuu vahasta tai muista polymeereistä. Kutikula toimii vesieristeenä ja auttaa estämään liiallista veden haihtumista sekä myös suojaa jossain määrin mikrobien hyökkäyksiltä. Useimmilla kasveilla on myös erikoistuneita pintahuokosia, ilmarakoja, joiden kautta kasvi voi ottaa ilmakehästä hiilidioksidia fotosynteesiin ja päästää ilmaan sen tuottamaa happea. Ilmaraot ovat myös väylä veden haihtumiselle kasvista; kuumissa ja kuivissa olosuhteissa ilmaraot sulkeutuvat ja minimoivat veden poistumisen.
Varhaisimmilta maakasveilta oikeat juuret ja lehdet puuttuivat. Fossiilit 420 miljoonan vuoden takaa paljastavat adaptaation, joka on voinut auttaa ravinteiden saantia: kasvit liittoutuivat sienien kanssa, aloittivat symbioosin niiden kanssa, ja näin kehitys kohti sienijuuria eli mykoritsoja sai alkunsa. Mykoritsasienet muodostavat valtavia rihmastoja maaperään, josta ne tuovat ravinteita kasvin käyttöön; vastineeksi ne saavat kasvilta yhteyttämistuotteita (sokereita). Symbioosi sienten kanssa mahdollisti jo juurettomien kasvien ravinteiden saannin.
Näkinparran muna- (ruskeat) ja siittiöpesäkkeitä (punaiset). – Christian Fischer [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)].Näkinpartaiset muistuttavat ulkonäöltään hieman kortteita, ja ennen niitä pidettiinkin vedenalaisina kortekasveina. Ne voi äkkiseltään sekoittaa myös eräisiin vitoihin tai hauroihin. Ne on aiemmin myös luokiteltu viherleviksi (Chlorophyta). Yhteistä viherlevien kanssa ovat pigmentit, erityisesti klorofylli a ja b, sekä vararavintotärkkelyksen kerääntyminen viherhiukkasiin. Ulkonäössä selvin ero useimpiin viherleviin on sekovarren jakaantuminen niveliin ja nivelväleihin. Lisääntymiselimet ovat monimutkaisia ja erikoisia, ja varsinkin koirasgameettien muoto osoittaa läheisempää sukulaisuutta sammaliin kuin viherleviin. Solun rakenne ja jakautuminen muistuttavat enemmän versokasveja kuin viherleviä. Charophyceae onkin osoittautunut Streptophyta-pääjaksoon kuuluvaksi luokaksi ja siten versokasvien (Embryophyta) sukulaiseksi. Näkinpartaisten luokassa on vain yksi lahko Charales, jossa yksi ainoa heimo Characeae, jossa kuutisen sukua, joissa yhteensä 80 lajia.
Streptofyytteihin kuuluva näkinpartaisten luokka (Charophyceae) oli monimuotoinen menneinä maailmankausina. Fossiileja on siluurikauden kerrostumissa, ja valtakausi lienee ollut jura- ja liitukaudella. Sekovarsiin ja varsinkin tsygootteihin (oospooreihin) kerääntyy kalkkia, joten jäänteitä eli ns. gyrogoniitteja on säilynyt.
RAKENNE ELI MORFOLOGIAA JA ANATOMIAA
Näkinpartaisten sekovarsi on pysty, ja siinä on haaroja kiehkuroina säännöllisin välein. Itäneessä taimessa joka toisesta solusta muodostuu nivelsolu ja joka toisesta pitkä nivelvälisolu. Nivelvälisolu voi olla jopa kymmenen sentin mittainen. Nivelsolu pysyy lyhyenä, ja pystysuuntaisten solunjakautumisten seurauksena syntyy rengas, jossa solujen lukumäärä on 6 – 20. Niiden jakautuessa syntyvät haarat ja näkinparroilla (suku Chara) myös kuorisolurihmat. Korvakekiehkurat ja gametangiot syntyvät myös nivelsolujen jakautumisen tuloksena.
Näkinpartaiset kiinnittyvät alustaan juurtumahapsien avulla, joita voi kehittyä myös ylempiin niveliin. Sekovarret voivat rönsyillä pitkin pohjaa. Eräillä lajeilla juurtumahapsiin syntyy talvehtimissilmuja eli bulbilleja.
Solunseinissä on selluloosaa, ja joissakin suvuissa solunseinän ulkokerroksissa on kalsiumkarbonaattia, joka tekee kasvin kovaksi ja hauraaksi. Soluissa on paljon viherhiukkasia. Pigmentteinä niissä on klorofylli a:ta ja b:tä kuten viherlevillä ja versokasveilla. Karotenoidejakin on, etenkin isolla punanäkinparralla (Charatomentosa). Soluissa on aluksi vain yksi tuma, mutta vanhoissa nivelvälisoluissa voi olla ehkä jopa yli tuhat pitkänomaista tumaa, jolloin tumat jakautuvat kuroutumalla. Kromosomit ovat melko kookkaita (3 – 5 mikronia eli mm: tuhannesosaa), ja ne muistuttavat ulkonäöltään esim. joidenkin liljakasvien kromosomeja. Solunjakautumisen yhteydessä syntyy fragmoplasti ja solulevy samantapaisesti kuin putkilokasveilla. Tuman jakautuessa tumajyvänen säilyy, mikä on alkukantainen ominaisuus; siemenkasveilla se häviää tumanjakautumisen aikana. Spermatogeneesin eli siittiösolujen synnyn yhteydessä on todettu sentrioleja, mutta niitä ei ole kasvullisten solujen jakautuessa.
lisääntyminen
Näkinpartaiset voivat lisääntyä suvuttomasti sekovarren palasista, joihin muodostuu juurtumahapsia eli ritsoideja. Palaset kulkeutuvat kauemmas, ja niistä kohoaa uusia pystyjä sekovarsia. Muuta suvutonta lisääntymistä ei ole.
Suvullinen lisääntyminen on oogamiaa. Lajit ovat joko yksi- tai kaksikotisia; edellisessä tapauksessa siittiö- ja munasolutuotanto tapahtuu samassa yksilössä, jälkimmäisessä eri yksilöissä (on erikseen koiraita ja naaraita). Gametangioiden eli sukusolupesäkkeiden rakenne on ainutlaatuinen. Näkinparroilla (Chara) ne syntyvät haarojen nivelkohtiin. Anteridiot eli siittiöpesäkkeet syntyvät värittömissä siittiöitö muodostavissa (spermatogeenisissa) rihmoissa eli anteridiorihmoissa. Rihmat ovat haarattomia, ja niissä on vain yksi solurivi. Kussakin laatikkomaisessa solussa syntyy yksi kaksisiimainen, pitkänomainen ja kierteinen siittiösolu eli spermatotsoidi. Se syntyy mitoosin tuloksena, sillä näkinpartaiskasvi on gametofyytti, jossa on yksiannoksinen kromosomisto (ts. se on haploidi). Ennen spermatogeenisten rihmojen syntyä solunjakautumisten tuloksena muodostuu kehittyviä siittiösoluja suojaava pallo, joka on läpimitaltaan 1 – 2 mm. Haaran nivelessä oleva kantasolu jakautuu kahdeksaksi primaariksi nuppisoluksi (kapitulum). Niiden jakautuessa syntyy pitkänomaisia kahvasoluja (manubrium). Ne suuntautuvat nuppisoluista ulos, ja niissä ovat litteät kilpisolut, jotka peittävät siittiöpalloa. Pallon keskellä olevat nuppisolut jakautuvat sekundaarisiksi nuppisoluiksi, joista syntyy spermatogeenisiä rihmoja. Yhtäläissekovartisilla lajeilla siittiöpallo sijaitsee nivelessä naarasgametangion eli oogonion alapuolella. Siittiöpallorakennetta kokonaisuudessaan ei voine tulkita anteridioksi eli koirasgametangioksi. Oogonion sisällä on suuri munasolu, ja oogoniota ympäröi viisi spiraalimaista, putkimaisten solujen muodostamaa rihmaa. Ne yhtyvät oogonion kärjessä koronaksi, jossa on joko viisi tai kymmenen solua. Kun siittiöt ovat valmiita, koiraspallojen kilpisolut irtoavat toisistaan, ja siittiösolut pääsevät ulos. Ne tunkeutuvat oogonion kruunusolujen tyvessä olevasta aukosta oogonioon, jossa munasolu hedelmöityy. Syntyy paksuseinäinen, ruskea tai musta lepovaiheinen tsygootti, ns. oospori. Siinä tapahtuu kaksi peräkkäistä jakoa, ja meioosi taphtunee tässä vaiheessa. Seuraa useita mitoottisia jakoja, ja nuori taimi tunkeutuu ulos tsygootin seinässä olevasta reiästä. Taimesta kehittyy uusi haploidi näkinpartainen. Vaikkakin näkinpartaisten kehityskierto on haplontti (so. haploidi vaihe eli gametofyytti hallitsee), esiintyy niillä kuitenkin polyploidiaa.
näkinpartaislajeja
Näkinpartojen sukuun, Chara, kuuluu kaikkiaan noin 20 lajia ja silopartojen sukuun, Nitella, noin 50 lajia. Kumpikin suku on levinnyt koko maapallolle. Laajalle levinnyt on myös Tolypella-suku, muut (Lamprothamnion, Lychnothamnus ja Nitellopsis) ovat vain 1 – 3 -lajisia ja harvinaisempia. Suomessa kasvaa parikymmentä näkinpartaislajia, joista joitakin seuraavassa. Monet lajit ovat maassamme suuria harvinaisuuksia ympäristövaatimustensa vuoksi. Esimerkiksi kalkkipitoisia järviä meillä on vähän.
Suurikokoisin Suomessa tavattava laji on punanäkinparta (Chara tomentosa), joka on väriltään useimmiten kellan- tai ruskeanpunainen. Se voi muodostaa laajoja kasvustoja merenlahtiin ja on tyypillinen pitkissä, kapeissa, melkein umpeen kuroutuneissa lahdissa. Esimerkiksi Tanskassa se kasvaa myös järvissä. Punanäkinparta on vankkarakenteinen, ja sillä on tavallisesti isot ja pulleat punertavat haarojen päätesolut sekä korvakkeet. Laji on kaksikotinen, mutta koirasyksilöt ovat Suomessa hyvin harvinaisia. Syksyllä sen alimpiin nivelsoluihin kertyy runsaasti tärkkelystä, ja kasvit talvehtivat jään alla. Pian jäiden lähdettyä lietteellä lojuvista edellisen vuoden päärangoista versoaa uusia päärankoja, joiden niveliin kehittyy nopeasti punakeltaisia, nuppineulan päätä muistuttavia siittiöpalloja. Siittiöiden valmistuttua pallot hajoavat. Repeytyneiden pallojen kilpisolut näkyvät usein liuskoina nivelkohdissa. Nuorten pallojen spermatogeeniset rihmat kasvavat hyvin nopeasti, ja solunjakautumisia on hyvin paljon. Ne sopivat kromosomitutkimukseen, sillä ne voivat olla jopa viiden mikronin pituisia ja värjäytyvät helposti tavallisilla värjäysmenetelmillä. Punanäkinparralla on 14 kromosomia (n=14). – kuvia
Otanäkinparta (Chara aspera) on noin 20 cm korkea, vihreä, piikkinen, ja kasvaa rannikoilla. Punanäkinparran tavoin se on kaksikotinen, ja kromosomilukukin on sama. Molemmat sukupuolet ovat yleisiä. Juurtumahapsissa on etenkin syksyllä yksisoluisia, valkoisia tärkkelyspallosia, joiden avulla laji talvehtii. Otanäkinparta on merialueittemme selvästi runsain ja koko rannikkoalueelle levinnyt. Juurtumahapsissa on pohjaliejun suojassa neljä valkeaa ja pyöreää talvehtimissilmua (bulbillia). – kuvia
Harvinaisempana otanäkinparran seurassa lounaassa suojaisissa merenlahdissa kasvaa samankokoinen tummanäkinparta (Chara canescens), jolla on värittömät kuorisolut lukuisine vihreine piikkisolurykelmineen. Väriltään se on tummanvihreä, kasvutavaltaan tiheähaarainen. Sekin on kaksikotinen, mutta maamme alueilla on vain naarasyksilöitä. – kuvia
Järvinäkinparta (Chara fragilis) kasvaa järvissä koko maassamme ja paikoin myös vähäsuolaisessa murtovedessä. Se on maassamme laajimmalle levinnyt näkinparta, ja sen löytää varmimmin järvenpohjan lähteiden tuntumasta. Se muistuttaa otanäkinpartaa, mutta on piikitön, vailla tärkkelyspallosia ja yksikotinen. Järvinäkinparta on ulkonäöltään vaatimaton, hento ja tummanvihreä. – kuvia
Yksikotinen on myös itämerennäkinparta (Chara baltica), joka on punanäkinparran kokoinen, yleensä hennompi, vihreä, haarova ja harvapiikkinen laji. Kummatkin kasvavat usein samantapaisilla kasvupaikoilla. Itämerennäkinparta on monivuotinen, ja sen pääranka ja haarat ovat uurteisia, mikä johtuu keskenään vuorottelevista eri-ikäisistä ja -kokoisista kuorisoluriveistä. – kuvia
Siloparroista (Nitella) Suomessa yleisimpänä esiintyy järvisiloparta (N. flexilis), joka nimensä mukaan on erityisesti järvien ja lintuvesien laji, jota kasvaa jokisuistojen ja rehevien järvien avoimissa osissa. Kooltaan se on suuri ja muistuttaa hapsivitaa (Potamogeton pectinatus). Se on kuoreton, ja sen päärangassa on senttimetrien pituisia nivelvälisoluja, joita on paljon käytetty solufysiologian tutkimuksissa. Tyypilliset, kertaalleen dikotomisesti eli halkihaaraisesti haaroittuneet haarat sekä gametangiot lähtevät nivelistä. – Karummissa järvissä kasvaa muita, pienempiä silopartoja. – kuvia
Tuulenpesäparta (Tolypella nidifica) on yksikotinen ja kasvaa rannikon murtovesialueella. Se on sileäkuorinen ja piikitön, heleänvihreä, ja sillä on yleensä runsaasti gametangioita, jotka ovat ryhmittyneet pesämäisiksi möykyiksi, mistä nimi johtuu. – kuvia
näkinpartaisten ekologiaa
Näkinpartaiset ovat kalkkialueiden kasveja. Niitä on monissa Itä-Suomen järvissä, eniten alueilla, missä on runsaasti järvikalkkia, esim. Kuusamossa. Lounais-Suomessa niitä on matalissa ja suojaisissa murtovesilahdissa, merenlahdista kuroutuneissa järvissä sekä vanhoissa kalkkilouhoksissa, joihin on kerääntynyt vettä.
Fosfaattien lisääntyminen eli vesien rehevöityminen, ruoppaus ja veneliikenne hävittävät näkinpartaiskasvustoja. Särkikalat käyttänevät ravinnokseen näkinpartaisia, sillä niitä löytyy kalojen ruoansulatuskanavasta. Myös joutsenet, hanhet, sorsat ja sotkat syövät mielellään näkinpartaisia.
Näkinpartaisilla on luonteenomainen, epämiellyttävä tuoksu, joka vaihtelee eri paikoilla ja eri aikoina. Se johtuu allelopaattisista eritteistä, joiden avulla kasvit ehkäisevät pintaa peittävien päällyslevien kiinnittymisen (kasvien kemiallista vaikuttamista toisiinsa sanotaan allelopatiaksi). Eritteidensä vuoksi näkinpartaisia on käytetty hyönteisten torjunnassa.
Lähteet
Kallio, P. & Rousi, A. (päätoim.) 1981: Kasvien maailma, Otavan iso kasvitietosanakirja, osa 4. Näkinpartaislevät. – Otava. ISBN 951-1-06068-6.
Vuokko, S. (päätoim.) 2001: Suomen luonto. Kasvit, osa 3; Bakteereja, sieniä, itiökasveja. – WSOY-yhtymä. ISBN 951-35-6614-5.
Streptofyyttien (pääjakso Streptophyta) kuutta ensimmäistä luokkaa pidettiin ennen viherlevinä (pääjakso Chlorophyta). Viherlevillä ja streptofyyteillä on yhteinen kantamuoto, ja jos mainitut luokat levämäisten ulkonäkönsä ja elintapojensa vuoksi sisällytettäisiin viherleviin, saataisiin ryhmä, joka ei ole monofyleettinen eli se ei sisältäisi kantamuotoa ja kaikkia sen jälkeläisiä. Siinä tapauksessa se olisi parafyleettinen. Nykyisessä systematiikassa tavoitellaan monofyleettisiä taksoneita.
Kuutta ensimmäistä luokkaa on myös kutsuttu yhteisnimellä Charophyta. Sitäkään ei voi monofyleettisyysvaatimusten mukaan pitää ns. oikeana taksonina, sillä sekin olisi parafyleettinen: se sisältäisi kantamuodon, muttei sen jälkeläisiä, joita nimitetään embryofyyteiksi eli versokasveiksi eli alkiollisiksi kasveiksi ja myös maakasveiksi varsinkin englanninkielessä (land plants).
Joka tapauksessa nuo kuusi luokkaa ovat versokasvien lähimpiä sukulaisia. Siitä on sekä morfologian että molekyylien antamia todisteita.
Versokasvit, jotka yleisesti miellämme varsinaisiksi kasveiksi – sammalet, sanikkaiset ja siemenkasvit – ovat monisoluisia eukaryootteja ja fotosynteettisiä autotrofeja eli omavaraisia eliöitä, joiden solunseinät ovat selluloosaa ja viherhiukkasten yhteyttämispigmentit klorofylliä a ja b. Vaikka monilla leväryhmillä on joitakin näistä ominaisuuksista, ei niistä millään ole näitä kaikkia. Esimerkiksi rusko-, puna- ja tietyt viherlevät ovat monisoluisia eukaryootteja ja yhteyttäviä. Selluloosaisia solunseinä on viher-, rusko- ja panssarilevillä, ja esim. viher- ja silmälevillä on samanlaiset klorofyllit kuin varsinaisilla kasveilla. Jo tästä vertailusta näkee, että viherlevät muistuttavat levistä eniten versokasveja.
Kaiken tämän lisäksi edellä mainituilla kuudella Charophyta-luokalla eli karofyyteillä on vielä lisää yhteisiä piirteitä versokasvien kanssa, mikä osoittaa, että ne ovat evoluution kulussa kehittyneet maakasveihin johtavasta linjasta ja ovat nykyään elävistä levistä niiden läheisimmät sukulaiset, vaikka ovat useimmiten yksisoluisia tai solukolonioita muodostavia. Näkinpartaiset (Charophyceae) ovat sekovartisia ja muistuttavat hieman ulkonäöltäänkin maakasveja – mutta vain pinnallisesti.
Sekä versokasveilla että karofyyteillä on luonteenomaisia rengasmaisia proteiineja plasmamembraanissaan eli solukalvossaan. Ne syntetoivat solunseinän selluloosakuidut. Muilla selluloosaseinäisillä levillä vastaavat proteiinit ovat lineaarisissa muodostelmissa.
Toiseksi, osalla kasveista on rakenteeltaan samankaltaisia uintisiimallisia siittiösoluja kuin karofyyteillä. Ja kolmas yhteneväisyys löytyy solunjakautumisesta, esim. rakenteesta nimeltään fragmoplasti.
Solun jakautuminen on tietyiltä osiltaan samanlainen versokasveilla ja karofyyteillä muttei muilla levillä. Fragmoplasti on mikrotubulusrykelmä, joka muodostuu jakautuvan solun tytärtumien väliin. Fragmoplastin keskelle muodostuu solulevy jakautuvan solun poikki. Se taas saa aikaan solunseinän muodostumisen, jolloin tytärsolut lopullisesti eroavat toisistaan.
Viherhiukkaset ja mitokondriot sisältävät solutuman lisäksi DNA:ta, mikä johtuu niiden endosymbionttisesta alkuperästä, kun syano- ja proteobakteeri ovat muuttuneet soluelimiksi. Näitä kutakin DNA:ta vertailemalla versokasvien ja karofyyttien välillä on saatu selville, että karofyyteistä yhtymälevät (Zygnematophyceae) ja Coleochaetophyceae ovat versokasvien lähimmät sukulaiset. Nykypäivän karofyytit voivat kertoa jotain versokasvien leväkantamuodoista, vaikka eivät itse olekaan niiden kantamuotoja, vaan samoista kantamuodoista kehittyneitä.
Vherhiukset eli vyörihmat (Ulothrix) ovat rihmamaisia viherleviä. – T.Voekler [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)].Viherlevät, Chlorophyta, ovat jo selvemmin kasvikuntaa kuin aikaisemmin esitellyt punalevät. Ne polvetuvat samasta kantamuodosta kuin ns. Streptophyta-ryhmä, johon kuuluu viherlevämäisiä sekä yksi- että monisoluisia muotoja (mm. näkinpartaiset) ja varsinaiset maaelämään sopeutuneet kasvit (sammalet, sanikkaiset, siemenkasvit). Viherlevistä ja streptofyyteistä käytetään yhteisnimitystä Viridiplantae (latinaa, ’vihreät kasvit’).
Viherlevien pääjaksoon, Chlorophyta, kuuluu yli 7000 lajia. Suurin osa niistä elää suolattomissa vesissä, vähemmistö merissä tai maalla.
Rakennetasot
Osa viherlevistä on yksisoluisia, ikäänkuin sukusoluasteelle jääneitä ajatellen monisoluisia lajeja (silti niilläkin on sukupolvenvuorottelua). Useat yksisoluisista elävät kuivalla maalla, mutta myös vesistöjen planktonissa. Yksisoluisista monet ovat myös symbioottisia muiden aitotumaisten kanssa, jolloin ne antavat yhteyttämistuotteitaan isäntänsä ravinnoksi. Jotkin kasvavat voimakasvaloisissa ja ultraviolettisäteilylle alttiissa ympäristöissä ja suojautuvat liialta paisteelta erinäisin yhdistein, joita on niiden solulimassa, solunseinässä tai tsygootin kuoressa; niiden väritys ei ole viherleville ominainen vihreä.
Viherleväsolut voivat järjestyä kolonioiksi, jotka ovat ovat tavallisesti rihmamaisia. Rihmalevät esiintyvät kelluvina tai pohjakasvillisuutena lammissa tai muussa matalassa vedessä. Esim. vyörihmat (Ulothrix – kuvia).
Todellisessa monisoluisessa rakennetasossa on solujen välillä erilaisuutta ja niillä erilaisia tehtäviä. Kasvu tapahtuu solujen lisääntyessä jakautumalla. Esimerkkeinä pallolevät (Volvox –kuvia) ja merisalaatit (Ulva – kuvia).
Solusulaumassa eli synsyyttisessä rakenteessa tapahtuu solun tumien jakautumisia ilman, että solut jakautuisivat. Esimerkiksi suikerolevien (Caulerpa) rihmamaiset haarat ovat solunseinättömiä ja monitumaisia synsyyttejä. – kuvia.
Lisääntyminen
Useimmilla viherlevillä on monimutkainen elämänkierto, jossa on sekä suvullisesti että suvuttomasti lisääntyviä vaiheita. Lähes kaikki lajit lisääntyvät suvullisesti kaksisiimaisten sukusolujen välityksellä, joissa on kuppimaiset viherhiukkaset eli kloroplastit.
Luokittelujärjestelmä
Viherlevien luokittelujärjestelmä näyttää nykyään tältä (Ruggiero, M.A., Gordon, D.P., Orrell, T.M., Bailly, N., Bourgoin, T., Brusca, R.C., Cavalier-Smith, T., Guiry, M. D. & Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms. PLoS ONE 10(4): e0119248.):
Pääjakso Chlorophyta
Alajakso 1. Chlorophytina. Viisi luokkaa, 23 lahkoa.
Luokka 1. Chlorodendrophyceae. Ainoastaan lahko Chlorodendrales.
Luokka 2. Chlorophyceae. Seitsemän lahkoa, joista yksi nimeämätön (siinä mm. Chlorangiopsidaceae-heimo). Muut lahkot: Chaetopeltidales, Chaetophorales, Chlamynomonadales (Volvocales), Oedogoniales ja Spheropleales.
Luokka 3. Pedinophyceae. Kolme lahkoa: Marsupiomonadales, Pedinomonadales ja Scourfieldiales.
Luokka 4. Trebouxiophyceae. Viisi lahkoa: Chlorellales, Microthamniales, Phyllosiphonales, Prasiolales, Trebouxiales.
Lahko Chlorodendrales (luokka Chlorodendrophyceae) sisältää tav. nelisiimaisia ja kuorellisia, muodoltaan monenmoisia yksisoluisia viherleviä. Kuori koostuu yhdistyneistä hiilihydraattipitoisista suomuista ja peittää yhtenäisenä koko soluruumiin; siimassa niitä on yksittäin. Suomujen ja kuooren hienorakenne vaihtlee lajista toiseen ja toimii tunnistamisen välineenä. Lajeja tavataan sekä meri- että sisävesissä keijustossa ja pohjaeliöstössä. Kaikki ovat yhteyttäviä ja omavaraisia (fotoautotrofeja). Useimmat elävät vapaina, eräät kuitenkin symbiontteina eläimissä. Suotuisissa olosuhteissa lajit voivat muodostaa leväkukintoja. – kuvia.
Lahkossa Chlamydomonadales eli Volvocales (luokka Chlorophyceae) on pitkä- tai lyhytsiimaisia viherleviä, jotka voivat muodostaa levymäisiä tai pallomaisia kolonioita, joissa on soluja 32 – yli 500 kpl. Kussakin solussa on kaksi siimaa, ja ne toimivat koordinoidusti. Suvuttomassa lisääntymisessä solut jakautuvat, kunnes uusi kolonia on syntynyt vapautettavaksi. Pienissä kolonioissa kaikki solut jakautuvat, suurissa on sekä kasvullisia että lisääntyviä soluja. Suvullinen lisääntyminen vaihtelee isogamiasta oogamiaan. Edellisessä sukusolut ovat samanlaisia keskenään ja liikkuvia, jälkimmäisessä munasolu on suuri ja liikkumaton. – kuvia.
Lahko Oedogoniales (luokka Chlorophyceae) käsittää suolattomien vesien rihmaleviä, ja on varsin selväpiirteinen ja omanlaisensa ryhmä. Esimerkiksi suvuttoman lisääntymisen liikkuvat itiöt (tsoospoorit) ovat monisiimaisia, ja siimat ovat järjestyneet kiehkuraksi solun kärjen tuntumaan. Suvullinen lisääntyminen on erikoislaatuista oogamiaa, ja solujen jakautumisessa on mutkikas menetelmä. Lahkoon kuuluu runsaat 400 lajia ja kolme sukua. – kuvia.
Sphaeropleales-lahkosta (luokka Chlorophyceae) on aiemmissa järjestelmissä käytetty nimeä Chlorococcales. Lahkoon kuuluu eräitä makean veden planktonin yleisimpiä leviä, kuten Scenedesmus ja Pediastrum. Lahkon levät ovat joko liikkumattomia yksisoluisia tai kolonioita muodostavia, joilla on kaksisiimaisia tsoospooreja. – kuvia.
Pedinomonadales (luokka Pedinophyceae) on yksiheimoinen lahko, jonka lajit ovat pieniä yksisoluisia leviä. Kussakin solussa on yksi siima.
Bryopsidales-lahkossa (luokka Ulvophyceae) sekovarsi on rihmamainen ja hyvin haarainen ja voi olla sotkeutunut massamaiseksi. Solut ovat monitumaisia eikä väliseiniä ole, joten kiekkomaiset kloroplastit ja muut soluelimet (organellit) voivat liikkua vapaasti kaikkialla sekovarressa. Koko levä voi olla yhtä ainutta solua, ja suikerolevillä (Caulerpa) se voi olla monta metriä. Halimeda-suvussa voi yksi ainoa yksilö muodostaa merenpohjaniityn, joka koostuu yhdestä solusulaumayksilöstä. Lahkon levien lisääntyminen on pääasiassa kasvullista: pienistä kappaleista kasvaa uusi yksilöitä, jolloin syntyy laajoja klooneja. Joissakin olosuhteissa esiintyy suvullista lisääntymistä, jossa sekovarren koko sytoplasma muuttuu kaksisiimaisiksi sukusoluiksi (ilmiötä nimitetään holokarpiaksi). Ne vapautuvat meriveteen solunseinän nystymäisistä aukoista, papilleista. Tsygootista eli hedelmöityneestä munasolusta kasvaa alkeisrihma (protoneema), joka kehittyy uudeksi yksilöksi.
Dasycladales (luokka Ulvophyceae) on viherlevälahko, joka koostuu kookkaista yksisoluisista lajeista (koko vaihtelee 2 – 200 mm). Ne elävät lämpimien merien matalissa osissa, tav. enintään 20 m:n syvyydessä. Suunnatonta solua peittää kalsiumkarbonaattinen kuori. Kasvullisessa vaiheessa niissä on vain yksi tuma. Vasta kun on aika tuottaa gameetteja, tuma jakautuu meioottisesti ja sen jälkeen lukuisia kertoja mitoottisesti suureksi määräksi haploideja tumia, jotka siirtyvät leväsolun yläosan gametangioihin (sukusolupesäkkeisiin). Huomattavaa solun rakenteessa on proteiinirihmojen muodostama verkosto. Lahkoon kuuluu mm. suku Acetabularia.
Lakkisienimäisiä Acetabularia-leviä meren pohjalla. – Tigerente [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)].Mamiellales-viherlevien (luokka Mamiellophyceae, alajakso Prasinophytina) solut ja siimat ovat yleensä seittimäisten suomujen peittämiä. Yhteistä suomuisille ja suomuttomille lajeille ovat tietyt pigmentit.
Micromonas pusilla, lahko Mamiellales. – No machine-readable author provided. Esv assumed (based on copyright claims). [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)].Palmophyllales (luokka Pyramimonadophyceae) ovat syvänmeren viherleviä. Viherleviksi ne ovat omalaatuisesti monisoluisia siten, että niiden solut sijaitsevat hyytelömäisessä väliaineessa. Sukuja lahkossa on vain kolme: Palmophyllum, Verdigellas ja Palmoclathrus. – kuvia
Suomen viherleviä
Maamme vesialueilla elää joitakin makroskooppisia viherlevälajeja, joista seuraavassa:
Salaattilevä, Monostroma grevillei, on arktisten seutujen levä, joka on jäänyt Itäereen jäänteenä jääkauden merivaiheista. Se on läpikuultava ja vihreä ja parhaiten havaittavissa jäiden lähdön aikoihin. Salaattilevä on sopeutunut alle neljäasteiseen, sulamisvesien viilentämään murtoveteen. Kasvu alkaa jo jään alla, ja kun veden lämpötila nousee yli kahdeksan asteen, sen elintoiminnot häiriintyvät. Kesät se viettää syvissä ja viileissä vesissä. – kuvia
Ahdinparta, Cladophora glomerata, muodostaa rannikoittemme kivikkoihin ja kallioihin jopa useita metrejä leveitä vihreitä vyöhykkeitä vesirajasta jopa viiden metrin syvyyteen. Se tuottaa eläimille paljon ravintoa ja toimii hyvänä lisääntymissuojana. Monet äyriäisten nuoruusvaiheet syövät ahdinpartaa ja sillä kasvavia piileviä sekä pienempiä rihmamaisia viherleviä ja monet mikroskooppiset ja vähän isommatkin selkärangattomat, kuten kotiloiden ja simpukoiden toukat, etsivät suojaa niitä saalistavilta kaloilta ja niiden poikasilta. Ahdinparta kasvaa runsaana myös makeissa vesissä, ja sen runsastuminen on vesien rehevöitymisen seurausta. – kuvia
Ahdinparralla on maassamme ainakin kaksi sukulaista. Kallioahdinparta, C. rupestris, kasvaa ulkosaariston tyrskyisillä kalliorannoilla Porin korkeudelle saakka. Se on väriltään oliivinvihreä, ahdinparta on kirkkaamman vihreä. Vasta lokakuussa kallioahdinparta saavuttaa näyttävimmän kasvustokokonsa. – kuvia. Palloahdinpartaa, C. aegagrophila, voi nähdä toukokuussa murtovedessä ja kirkkaissa järvissä tummanvihreinä ja takkuisina kelluvina palloina, joiden läpimitta on kaksi – kolme senttiä. Tavallisesti se muodostaa pohjalle mattomaisia kasvustoja. – kuvia.
Suolilevä, Enteromorpha intestinalis, on nimensä mukaisesti suolenpätkän kaltainen putki ulkosaariston luotojen rehevissä kalliolammissa. Niissä se on ainoa muutaman sentin mittaiseksi kasvava kasvi. Kalliolammikoiden lisäksi sen voi yksittäisinä löytää ahdinpartavyöhykkeestä, laitureilta ja kiviltä vesirajassa. – kuvia.
Aitotumaisten kolmatta pääryhmää voisi kutsua kasvikunnaksi sanan laajimmassa merkityksessä. Useinhan on vain mielipidekysymys, mihin kohtaan fylogeneettistä kaavaa eli evoluutiopuuta mikäkin termi sijoitetaan. Yleensä kuitenkin kasvikunta määritellään ahtaammin, joten käytän tässä ryhmän tieteellistä nimeä Archaeplastida.
Systemaattinen asema eliökunnassa
Esitumaiset, Prokaryota
Aitotumaiset, Eukarya
Ekskavaatit, Excavata
SAR
Archaeplastida
Punalevät, Rhodophyta
Glaucophyta
Viherlevät, Chlorophyta
Kasvit, Streptophyta
Yksisiimaiset, Unikonta
Punaleviä: A-D Chondrus crispus -lajin erilaisia muotoja, E-F Gigartina mamillosa eli Mastocarpus stellatus. – Franz Eugen Köhler, Köhler’s Medizinal-Pflanzen [Public domain].Archaeplastida-ryhmässä endosymbionttinen syanobakteeri plastidi-nimisenä soluelimenä on sääntö. Plastidien syntymistä endosymbioosin seurauksena tukevat sekä morfologiset että molekyylitodisteet. Puna- ja viherlevät ovat polveutuneet kantamuodosta, jossa kyseinen endosymbioosi ja sen seurauksena fotosynteesi syntyivät, ja viherlevämäisistä kantamuodoista kehittyivät kasvit. Archaeplastida on siten monofyleettinen ryhmä, joka sisältää kantamuodon ja kaikki sen jälkeläiset.
Punalevät
Punaleviä, Rhodophyta (tieteellinen nimi tulee kreikan sanoista rhodos ’punainen’ ja fyton ’kasvi’), tunnetaan noin 6000 lajia, ja useimmat ovat monisoluisia ja makroskooppisia, paljain silmin havaittavia. Niiden yleisväritys on punertava, mikä johtuu fotosyntetoivasta pigmentistä nimeltä fykoerytriini, joka peittää alleen vihreän klorofyllin. Mitä syvemmällä punalevä kasvaa sen punaisempi ja tummempi se on; aivan rantavedessä kasvavat ovat melko vihreitä. Lisäksi on lajeja, jotka ovat väriaineettomia ja elävät loisina toisilla punalevillä.
Runsaimmillaan punalevät ovat trooppisten merien rannikkovesissä. Eräät niiden pigmenteistä (mm. fykoerytriini) pystyvät absorboimaan sinisiä ja vihreitä aallonpituuksia, jotka tunkeutuvat suhteellisen syvälle veteen. Eräs punalevälaji on löydetty Bahamasaarten vesistä yli 260 m:n syvyydestä. Eräät lajit kasvavat makeassa vedessä ja eräät maalla.
Punalevien plastidit ovat kaksikalvoisia. Sisempi on peräisin endosymbiontista ja ulompi punaleväsolun vakuolikalvosta. Aivan kuten syanobakteereilla on plastideissakin fykobiliinejä (esim. fykoerytriiniä), jotka absorboivat yhteyttämisessä tarvittavaa valoenergiaa. Niiden lisäksi plastideissa on klorofylli a:ta ja karotenoideja (keltaisia väriaineita). Yksisoluisilla on vain yksi, enemmän tai vähemmän liuskoittunut plastidi, monisoluisilla puolestaan tavallisesti monta kiekkomaista. Yhteyttämisen tuloksena syntyneet hiilihydraatit varastoituvat floriditärkkelyksenä plastidien ulkopuolelle.
Punalevien solunseinä on yleensä kaksikerroksinen. Sisempänä on pääasiassa selluloosasta koostuva kerros, ulompana paksu ja löyhä, hyytelömäinen ja punaleville ominainen polysakkaridikerros. Monilla lajeilla solunseinään kasautuu myös kalkkia (erityisesti korallilevillä, Corallinales-lahko). Monisoluisilla on tavallisesti solunseinissä tulpan sulkemia huokosia eli kuoppakytköksiä. Tulpan hävitessä mahdollistuu aineiden siirtyminen solusta toiseen tarvittaessa.
Monisoluisilla punalevillä on alustaan kiinnittynyt sekovarsi, joka on tavallisesti melko pieni, enimmillään noin metrin mittainen. Sekovarsi koostuu toisiinsa kietoutuneista rihmoista, ns. pseudoparenkyymisolukosta (parenkyymi on kasvien perussolukko; pseudo viittaa epäaitouteen). Hyytelömäiset polysakkaridit sitovat solukoita yhteen. Sekovarsi kasvaa rihmojen kärkisolujen jakautuessa.
Punalevät lisääntyvät seksuaalisesti, ja niillä on monenlaisia elinkiertoja, joihin kuuluu yleisesti myös suvuttomasti lisääntyvä sukupolvi (sukupolvenvuorottelu). Gameetit eli sukusolut eivät ole siimallisia ja liikuntakykyisiä, niiden kohtaaminen on veden virtausten varassa. Tyypillinen elinkierto on kolmivaiheinen. Sukusoluja tuottava gametofyytti on haploidi eli kromosomistoltaan yksiannoksinen. Diploidi sukupolvi on kaksivaiheinen: karposporofyytti tuottaa diploideja itiöitä, tetrasporofyytti haploideja itiöitä. Gametofyytti ja tetrasporofyytti elävät itsenäisesti, karposporofyytti sen sijaan kiinni gametofyytissä ja on siitä riippuvainen. Tavallisesti gametofyytit ja tetrasporofyytit ovat ulkoisesti samanlaisia.
Punaleviä on ollut maapallon merissä jo kambrikaudelta lähtien melkein 600 miljoonan vuoden ajan.
Suomen punaleviä
Punalevät muodostavat myös Suomen rannikoilla alimman levävyöhykkeen noin 10-20 metrin syvyydessä. Muutamat valtamerilajit kasvavat meidänkin merialueiden syvänteissä, missä vesi on hieman suolaisempaa kuin pinnalla. Meillä ne eivät lisäänny suvullisesti ja jäävät pieniksi, vain muutaman sentin kokoisiksi.
Rannikoittemme yleisin punalevä on punahelmilevä (Ceramium tenuicorne; kuvia), jonka nimi johtuu varren tiheistä pyöreistä haarakimpuista, joten levä on helminauhamainen. Syksyisin sitä voi nähdä ajautuneena rannoille. Se kasvaa uloimpien järviruokojen varsilla, laituripaaluilla, merimerkeillä ja ulkosaariston kalliorannoilla.
Hildenbrandia prototypus (kuvia) muodostaa ohuen punaisen kerroksen usean metrin syvyydessä kivien pinnalle. Sen voi löytää lähempää rantaakin, kun jäät tai aallokko pitävät kookkaammat levät poissa.
Muutama punalevälaji kasvaa makeassa vedessä. Koskipunalevä (Lemaneafluviatile; kuvia) kasvaa ruskeanvihreinä, jäykkinä ja nivelikkäinä rihmoina koskien kivillä. Helminauhaleviä (Batrachospermum) löytää puolestaan lammista, missä ne kasvavat risuihin, korsiin ja kiviin kiinnittyneinä. Ne muodostavat muutaman sentin mittaisia limaisia ja sinivihreitä tupsuja ja ovat sopeutuneet kirkkaaseen valoon (kuvia)
Lisää kotimaamme punaleviä löytyy tästä (Luontoportti).
Punalevätuotteita
Punalevistä saadaan monia arkipäiväisiä tuotteita. Suuri osa punalevien solunseinästä on hyytelömäistä ainetta, joka liukenee kuumaan veteen. Teollisesti tuotettuna se tunnetaan nimellä agar-agar tai karrageeni. Niitä käytetään elintarvike-. lääke-, kosmetiikka- ja kemianteollisuudessa, ja niitä kerätään niihin tarpeisiin jo Tanskan ja Norjan rannikoilla. Samaan tapaan käytetään alginaatteja, joita saadaan ruskolevistä (Phaeophyta).
Japanissa punaleviä viljellään rannikkovesissä köysissä ja verkoissa. Niitä käytetään ravinnoksi tuotenimillä dulse ja nori. Jälkimmäistä käytetään mm. sushien päällimmäisenä kääreenä.
Punalevien luokittelu
Punalevien pääjakso, Rhodophyta, käsittää neljä alajaksoa, joissa on yhteensä seitsemän luokkaa, tässä lähteenä käytetyn, vuodelta 2015 peräisin olevan luokittelun mukaan.
Lähde: Ruggiero, M.A., Gordon, D.P., Orrell, T.M., Bailly, N., Bourgoin, T., Brusca, R.C., Cavalier-Smith, T., Guiry, M. D. & Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms. PLoS ONE 10(4): e0119248
Alajakso 1. Cyanidiophytina
Luokka Cyanidiophyceae: yksi lahko (Cyanidiales)
Alajakso 2. Eurhodophytina
Luokka Bangiophyceae: kaksi lahkoa (Bangiales, Goniotrichales)
Luokka Florideophyceae
Alaluokka tuntematon: lahko Rhodachlyales
Alaluokka Ahnfeltiophycidae: kaksi lahkoa (Ahnfeltiales, Pihiellales)
Alaluokka Corallinophycidae: kolme lahkoa (Corallinales, Rhodogorgonales, Sporolithales)
Alaluokka Hildenbrandiophycidae: yksi lahko (Hildenbrandiales)
Luokka Compsopogonophyceae: kolme lahkoa (Compsopogonales, Erythropeltidales, Rhodochaetales)
Alajakso 4. Rhodellophytina
Luokka Porphyridiophyceae: yksi lahko (Porphyridiales)
Luokka Rhodellophyceae: kolme lahkoa (Dixoniellales, Glaucosphaerales, Rhodellales)
Luokka Stylonematophyceae: kaksi lahkoa (Rufusiales, Stylonematales)
Glaukofyytit
Glaukofyytit (Glaucophyta, kuvia ) ovat punalevien yksisoluisia sukulaisia. Ne ovat vesieliöitä, joita tavataan lähinnä suolattomista pintavesistä. Joillakin lajeilla on kaksi sileää uintisiimaa, mutta toisilta ne ovat hyvin surkastuneita. Glaukofyyttien plastideja nimitetään syanelleiksi, sillä ne eroavat monessa suhteessa muista plastideista. Joissakin on esim. syanobakteereille tyypillinen, peptidoglykaania sisältävä solunseinä, vaikka niiden endosymbioottiset syanobakteerit ovat muutoin pitkälle surkastuneita.
Glaukofyyttejä tunnetaan vain runsaat kymmenen lajia, eivätkä ne ole erityisen yleisiä missään. Lajit ryhmitellään viideksi suvuksi: Cyanophora, Cyanoptyche, Gloeochaete, Glaucocystopsis ja Glaucocystis.
Cercomonas sp. -alkueliö eli protisti edustaa Cercozoa-pääjaksoa. – ja:User:NEON / commons:User:NEON_ja [CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5)].Cercozoa-pääjakso on muodostettu hyvin monenlaisista aitotumaisista yksisoluisista ns. alkueläimistä molekyylianalyysien perusteella. Morfologisesta moninaisuudestaan huolimatta ryhmää pitävät koossa ribosomien RNA:n ja eräiden muiden molekyylien samankaltaisuus, jonka perusteella ne ovat yhtä ja samaa evoluutiopuun haaraa.
Systemaattinen asema eliökunnassa:
Esitumaiset, Prokaryota
Aitotumaiset, Eukarya
Ekskavaatit, Excavata
SAR
Stramenopila (mm. pii- ja ruskolevät)
Alveolata (mm. itiöeliöt)
Rhizaria (valejalkaisia eliöitä)
sädejalkaiset, Radiolaria
huokoseliöt, Foraminifera
Cercozoa
Archaeplastida (mm. kasvit)
Unikonta, yksisiimaiset (mm. sienet ja eläimet)
Cercozoa kuuluu valejalkaisiin eliöihin, ja niillä on rihmamaiset valejalat ja uintisiimoja. Edelliset toimivat ravinnon pyydystäjinä, jälkimmäiset liikuntaeliminä.
Cercozoa-eliöitä tavataan yleisinä meri-, sisävesi- ja maaekosysteemeissä.
Valtaosa Cercozoa-pääjakson lajeista on toisenvaraisia eli heterotrofeja. Niissä on toisten yksisoluisten, kasvien ja eläinten loisia sekä petoja. Saalistuksen kohteina ovat enimmäkseen bakteerit, mutta myös toiset yksisoluiset, sienet ja jopa pienet eläimet. Ainakin yksi laji on omavarainen eli autotrofi, ja on myös miksotrofeja.
Eräs vähälajinen ryhmä, trooppisissa merissä tavattavat verkkolevät (Chlorarachnea) ovat ravitsemukseltaan miksotrofisia; so. ne pyydystävät bakteereita ja itseään pienempiä alkueliöitä eli protisteja sekä kykenevät myös yhteyttämään. Niiden endosymbioottinen yhteyttävä elin on alkuperältään viherlevä – ei syanobakteeri, kuten yleensä fotosynteettisillä eliöillä. Verkkolevien solut ovat amebamaisia, ja ne voivat muodostaa verkkomaisia limakkoja, jotka pystyvät liikkumaan hitaasti limasienten (Myxomycota) tavoin. Verkkolevät voivat tuottaa paksuseinäisiä lepoitiöitä, joista itää suotuisissa olosuhteissa uusia amebamaisia soluja tai parveiluitiöitä, jotka liikkuvat nopeasti siimansa avulla.
Eräs verkkolevä, Chlorarachnionreptans. – ja:User:NEON / commons:User:NEON_ja [CC BY-SA 2.5 (https://creativecommons.org/licenses/by-sa/2.5)].Paulinella chromatophora on ainut tunnettu omavarainen Cercozoa-ryhmän laji, ja sen solussa on suuri makkaramainen kromatofori, joka yhteyttää hiilidioksidia ja vettä auringonvalon avulla. Sen kromatoforin rakenne osoittaa, että se on alkuperältään endosymbionttinen syanobakteeri, kuitenkin eri syanobakteeri kuin varsinaisia viherhiukkasia eli kloroplasteja muodostava. – kuvia
Seuraavassa Cercozoa-pääjakson luokittelu vuodelta 2015 (Ruggiero ja muut 2015). Alajaksoja on kolme, luokkia yhteensä 11.
Alajakso Endomyxa
Luokka Ascetosporea: neljä lahkoa (Claustrosporida, Haplosporida, Paradinida ja Paramyxida)
Luokka Gromiidea: kaksi lahkoa (Gromiida ja Reticulosida)
Luokka Phytomyxea: kaksi lahkoa (Phagomyxida ja Plasmodiophorida)
Luokka Vampyrellidea: yksi lahko (Vampyrellida)
Alajakso Monadofilosa
Luokka Imbricatea
Alaluokka Placonuda: viisi lahkoa (Discocelida, Discomonadida, Euglyphida, Marimonadida ja Variglissida)
Alaluokka Placoperla: viisi lahkoa (Perlofilida, Rotosphaerida, Spongomonadida, Thaumatomonadida ja Zoelucasida)
Luokka Metromonadea: kaksi lahkoa (Metopiida ja Metromonadida)
Luokka Sarcomonadea: viisi lahkoa (Cercomonadida, Glissomonadida, Pansomonadida, Pseudosporida ja Sainouroida)
Luokka Thecofilosea
Alaluokka Eothecia: neljä lahkoa (Cryomonadida, Ebriida, Matazida, Ventricleftida)
Alaluokka Phaeodaria: kaksi lahkoa (Eodarida, Opaloconchida)
Alaluokka Tectosia: yksi lahko (Tectofilosida)
Alajakso Reticulofilosa
Luokka Chlorarachnea, verkkolevät: yksi lahko (Chlorarachnida)
Luokka Granofilosea: neljä lahkoa (Cryptofilida, Desmothoracida, Leucodictyida, Limnofilida)
Luokka Sciomonadea: yksi lahko (Tremulida)
Luokan Ascetosporea lajit ovat eläinten, erityisesti merten selkärangattomien, loisia, jotka tuottavat rakenteeltaan yksinkertaisia itiöitä. Haplosporida-lahkossa niissä on yksi ainoa tuma, lahkon Paramyxida edustajilla itiöt ovat monitumaisia.
Luokan Gromiidea eliöt muistuttavat ulkoiselta olemukseltaan verkkoleviä (Chlorarachnea).
Keskellä verkkolevämäinen Gromia, (luokka Gromiidea) laidoilla huokoseliötä (Foraminifera). – various [Public domain].
Luokka Phytomyxea (fytomyksidit) sisältää kasvien loisia ja aiheuttavat niissä äkämämäisiä muodostumia. Huomattavimmat niiden aiheuttamat kasvisairaudet ova kaalin ja sen sukulaisten möhöjuuri eli kaalirevännäinen, jota aiheuttaa Plasmodiophorabrassicae (kuvia), ja perunarutto, jonka aiheuttaja on Phytophthora infestans (kuvia). Kasvullinen vaihe on monitumainen solu, jota nimitetään plasmodioksi. Se jakautuu lopulta ja muodostaa uusia itiöitä, jotka vapautuvat isännän solujen hajotessa. Eri vaiheissa muodostuu lepoitiöitä ja liikkuvia tsoospooreja. Jälkimmäisillä on kaksi siimaa. Plasmodion jakautuvat tumat ovat luonteenomaisen ristimäisiä muodoltaan.
Perinteisesti fytomyxideihin kuuluvan lahkon Plasmodiophorida lajeja on pidetty limasieninä, joita ne muistuttavat plasmodioltaan (em. kaalirevännäinen kuuluu tähän lahkoon). Genetiikan ja hienorakenteen tutkimukset ovat kuitenkin osoittaneet, että niiden paikka eliökunnan luokittelussa on Cercozoa-pääjakson fytomyksidien luokassa.
Luokka Vampyrellidea käsittää paljaspintaisia, kuorettomia yksisoluisia, joilla on rihmamaisia sädemäisiä valejalkoja ja oranssin värinen, pallomainen ja läpimitaltaan 30 mikronin suuruinen soluruumis; eliö muistuttaa sädejalkaisia (Radiolaria). Liikkuessaan soluruumis muuttuu amebamaisemmaksi. Ravintonaan vampyrellidit käyttävät leväsoluja tai ovat sienten loisia, jollaisina niillä voi olla merkitystä viljojen ruostesienten rajoittajina.Vampyrella lateritia -vampyellidi. – Giuseppe Vago [CC BY 2.0 (https://creativecommons.org/licenses/by/2.0)]
Monadofilosa-alajakson lajit ovat amebamaisia, joko kuorellisia tai kuorettomia. Niitä tavataan merissä, makeissa vesissä ja sammalikoissa. Osa on siimallisia, ja ne liikkuvat etupään värekarvojen ja/tai rihmamaisten pseudopodien avulla. Luokkia alajaksossa on neljä. Imbricatea-luokkaan kuuluvilla on pinnallaan piipitoisia suomuja.
Alajaksoon Reticulofilosa kuuluvista verkkolevistä (luokka Chlorarachnea) on jo ylempänä ollut puhetta. Luokkaan Granofilosea luetaan eräitä ennen ns. aurinkoeläiminä (Heliozoa; kuvia) pidettyjä ryhmiä, kuten mm. lahko Desmothoracida. Kolmas alajakson luokka on nimeltään Sciomonadea.
Meren pehjassa eläviä huokoseliöitä. – This image is in the public domain in the United States because it only contains materials that originally came from the United States Geological Survey, an agency of the United States Department of the Interior. For more information, see the official USGS copyright policy..
Huokoseliöitäkin (Foraminifera) pidettiin aiemmin yksisoluisina alkueläiminä; siitä nimitys huokoseläimet. Niiden muodostaman pääjakson sekä suomalainen että tieteellinen nimi johtuu niiden soluruumista peittävästä kuoresta (testa), joka on täynnä huokosia (lat. foramen; ferre ’kantaa, kuljettaa’). Kuori on yksiosainen, mutta usein monilokeroinen, ja kalsiumkarbonaatin vahvistama. Outoa kyllä yksisoluisten kyseessä ollen, mutta suurimmat huokoseläinkuoret ovat halkaisijaltaan useita senttimetrejä.
Valejalat pistävät ulos kuoren huokosista ja toimivat uintieliminä, kuoren rakentajina ja ravinnon hankkijoina. Monet huokoseliöt saavat ravintoa myös symbionttisten levien fotosynteesistä.
Huokoseliöitä tavataan sekä merissä että sisävesissä. Enin osa elää hiekanjyvien seassa tai kiinnittyneenä kiviin tai leviin. On myös planktisia lajeja.
Kaikista tunnetuista huokoseliölajeista 90 % on fossiilisia. Yhdessä muiden kalkkipitoisten eliöiden kanssa huokoseliöt muodostavat merien pohjaan kerrostumia. Muinaisista pohjakerrostumista on muodostunut myös kallioita, jotka nykyään ovat kuivaa maata. Niiden ikää voidaan päätellä niiden sisältämistä huokoseliöfossiileista, ja eri puolilla maapalloa olevat muodostumat voidaan osoittaa samanikäisiksi. Ne paljastavat myös ilmastossa tapahtuneita muutoksia ja niiden vaikutuksia meriin ja niiden elämään.
Erään viimeaikaisen luokittelun mukaan (Mikhalevich 2013) huokoseliöiden pääjakso käsittää kuusi luokkaa, joista useimmissa on useita lahkoja. Lahko Reticulomyxida on systemaattiselta asemaltaan epävarma, eikä sitä ole kyetty sijoittamaan mihinkään luokkaan. Edellä mainittu luokittelu käsittää seuraavat luokat ja lahkot (alaluokat, ylälahkot ym:t välikategoriat on tässä jätetty huomiotta):
Luokka 1. Schizocladea, ainoa lahko Schizocladida.
Luokka 2. Xenophyophorea. Tärkeitä merien pohjaeliöitä, joita tavataan kaikissa valtamerissä mannerjalustoilta syvänteisiin; solu monitumainen; kuori kooltaan muutamasta millimetristä jopa 20 senttimetriin, muodoltaan vaihteleva, joskus rakenteeltaan monimutkainen; monimuotoinen luokka. Kaksi lahkoa: Stannomida ja Psamminida. – kuvia
Luokka 3. Astrorhizata. Kuori yksilokeroinen ja koostuu yhteen liimautuneesta materiaalista tai kalkista; muodoltaan pallomaisia, pitkänomaisia tai epäsäännöllisiä. Kahdeksan lahkoa: Ammoscalariida, Lagynida, Allogromiida, Astrorhisida, Dendrophryida, Hippocrepinida, Parathuramminida (kuollut sukupuuttoon), Psammosphaerida. – Lahkon Allogrimiida lajien kuori on enimmäkseen orgaanista ainesta, muodoltaan yksinkertainen, yksikammioinen; sekä mereisiä että sisävesilajeja; varhaisimmat huokoseliöfossiilit kuuluvat tähän lahkoon. – kuvia
Luokka 4. Rotaliata. Kuori moniosainen, enimmäkseen spiraalimainen; siinä yksi tai useampia aukkoja; kuoren sisäosa voi olla monimutkainen sekundaarisine kammioineen ja niitä yhdistävine kanavineen; kuori koostuu lasimaisesta kalsiitista tai yhteen liimautuneesta materiaalista. 24 lahkoa: Cassigerinellida, Globigerinida, Hantkenidia, Heterochelicida, Globorotaliida, Nautiloculinida, Spiroplectamminida, Textulariida, Trochamminida, Verneulinida, Robertinida, Elphidiida, Nummulitida, Orbitoidida (sukupuuttoon kuollut), Nonionida, Cassidulinida, Buliminida, Bolivinitida, Chilostomellida, Discorbida, Glabratellida, Planorbulinida, Rotaliida ja Rosalinida. – kuvia
Lahko Globigerinida. Merten planktonissa; kuori lasimaista kalsiittia, muodoltaan spiraalimainen, pitkäpiikkinen; kuori muodostaa pallomaisia kammioita ja tavallisesti siinä on yksi aukko, joskus useampia; fossiileja jurakaudelta lähtien; lajeja tunnetaan yhteensä yli 400, joista nykyään eläviä noin 30; tunneuin suku on Globigerina, jonka aiheuttamaa liejua on laajoilla alueilla merien pohjalla. – kuvia
Lahko Textulariida. Kuori muodostuu yhteen liimautuneesta ja kalkkisementin sitomasta materiaalista; fossiileja kambrikaudelta lähtien. – kuvia
Lahko Trochamminida. Fossiileja kambrikaudelta lähtien. – kuvia
Lahko Robertinida. Fossiileja keski-triaskaudelta lähtien; merten pohjaeliöitä; kuori spiraalimainen, monikammioinen, lasimainen. – kuvia
Lahko Buliminida. Enimmäkseen merenpohjalla; kuori huokoista, lasimaista kalsiittia; kammiot kaden tai kolmen sarjoissa ainakin varhaisvaiheessa. – kuvia
Lahko Rotaliida. Enimmäkseen valtamerten pohjaeliöstössä, eräät lajit jokisuistoissa; kuori lasimaista kalsiittia, monilokeroinen, tyypillisesti kierteinen; kammiot voivat olla sekundaaristen väliseinien jakamia; kuoren pinta voi olla eri tavoin nystyinen tai harjuinen; aukko samoin varustukseltaan vaihteleva. Lahkoon kuuluu paljon tärkeitä fossiileja, kuten nummuliitit. – kuvia
Lahko Lagenida. Merten pohjaeliöstöä; kuori kalsiittia, kammiot peräkkäin 1-3 sarjassa; kuoren muodot eräillä lajeilla viuhkamaisia tai lehtimäisiä; fossiileja ylä-siluurikaudelta lähtien. – kuvia
Luokka 6. Spirillinata. Spiraalimaisia, kuori joko yhteen liimautunutta materiaalia tai kalkkia. Lahkot: Plagioraphida (sukupuuttoon kuollut), Ammodiscida, Ammovertellinida, Ataxophragmiida, Archaediscida (sukupuuttoon kuollut), Lasiodiscida (sukupuuttoon kuollut), Tetradaxida (sukupuuttoon kuollut), Hottingerellida (sukupuuttoon kuollut), Involutinida (fossiileja varhais-permikaudelta myöhäis-liitukaudelle), Seabrookiida, Cymbaloporida, Spirillinida (kuori putkimainen ja kiertynyt, kalsiitista muodostunut; sukusolut eli gameetit ovat amebamaisia, kun taas muilla huokoseliöillä ne ovat kaksisiimaisia), Patellinida, Miliolata (muodoltaan vaihtelevia; kuori posliinimainen, joillakin siihen liittyy ympäristöstä peräisin olevia aineksia), Lituotubida, Loftusiida, Sphaeramminida, Cyclolinida, Haplophragmiida, Schlumbergerinida (eli Rzehakinida), Lituolida (fossiileja kambrikaudelta alkaen), Ozawainellida (sukupuuttoon kuollut), Endothyroida (sukupuuttoon kuollut), Tournayellida (sukupuuttoon kuollut), Fusulinida (sukupuuttoon kuollut; ala-siluurikaudelta ylä-permikaudelle; lajit ovat erinomaisia johtofossiileja kerrostumien ikää määritettäessä; paikoin niiden kuoret ovat merkittävä osa kalkkikiveä), Neoschwagerinida (sukupuuttoon kuollut), Schubertellida (sukupuuttoon kuollut), Schwagerinida (sukupuuttoon kuollut), Staffellida (sukupuuttoon kuollut), Costiferida (sukupuuttoon kuollut), Squamulinida, Cornuspirida, Soritida, Nebeculariida, Miliolida (kalkkikuori posliinimainen, sisäosa tavallisesti pseudokitiiniä; kuori magnesiumpitoinen ja orgaanista ainesta sisältävä, huokoseton ja monikammioinen; fosiileja kivihiilikaudelta alkaen; merten pohjaeliöstöä rannikoilta syvänteisiin). – kuvia
Ryhmä nimeltä Rhizaria käsittää sikäli amebamaisia eliöitä, että niiden edustajat liikkuvat ja hankkivat ravintoa ns. valejalkojen eli pseudopodien avulla. Pseudopodit ovat solun ulokkeita, jotka voivat vaihtaa muotoaan ja tulla esiin lähes mistä tahansa solun pinnan kohdasta. Ameba ankkuroi sen kärjen johonkin kohtaan ja solulima alkaa valua sitä kohden.
Rhizaria-ryhmän valejalkaiset eroavat tyypillisestä ameba-mallista morfologisesti. Näiden pseudopodit ovat rihmamaisia, ja lisäksi ryhmään kuuluu muotoja, joilla rihmamaisten pseudopodien ohella on uintisiima. Edellisiä ne käyttävät ravinnon hankkimiseen, jälkimmäistä liikkumiseen.
Tämän artikkelin eliöryhmän systemaattinen asema eliökunnassa:
Eliökunta
Esitumaiset eli tumattomat (Prokarya)
Aitotumaiset eli tumalliset (Eukarya)
Ekskavaatit (Excavata)
SAR
Sukasiimaiset (Stramenopila)
Alveolaatit (Alveolata)
Valejalkaiset (Rhizaria)
Sädejalkaiset (Radiolaria)
Huokoseliöt (Foraminifera)
Cercozoa
Archaeplastida (mm. kasvit)
Unikonta (mm. sienet ja eläimet)
Sädejalkaisia. – Luis Fernández García. 2007-03-26 This file is licensed under the Creative Commons Attribution-Share Alike 2.5 Spain license.
Sädejalkaiset
Sädejalkaisten (Radiolaria) solua tukee sisäinen tukiranka, joka on rakentunut pääasiassa piistä ja on rakenteeltaan usein hyvinkin kauniin symmetrinen. Ryhmän lajit elävät pääosin merivedessä planktisina. Pseudopodit lähtevät säteittäisesti solun keskiosasta, ja niiden tukena on mikrotubuluskimppuja, joiden päällä on ohut kerros solulimaa eli sytoplasmaa. Se nielee pienet mikroeliöt, jotka tarttuvat valejalkoihin, ja sen virtaus kuljettaa ravinnon solun muihin osiin. Tämänlaisia pseudopodeja sanotaan aksopodeiksi. Sädejalkaisten kuoret vaipuvat eliöiden kuoltua merenpohjaan, jossa ne muodostavat paikoin jopa satojen metrien paksuisia lietekerrostumia.
Sädejalkaisen solu on läpimitaltaan 0.1 – 0.2 mm. Sisäinen tukiranka jakaa solun endo- ja ektoplasmaksi kutsuttuihin sisä- ja ulko-osaan. Tuma ja muut soluelimet eli organellit sijaitsevat endoplasmassa. Ektoplasma sisältää rakkoja eli vakuoleja ja lipidipisaroita, jotka edistävät eliön kelluvuutta. Usein solussa on myös endosymbionttisia leväsoluja (erit. zooksantelli-panssarileviä), jotka tuottavat suurimman osan solun tarvitsemasta energiasta.
Tukirangan vuoksi sädejalkaiset fossiloituvat helposti, ja niitä tunnetaan kambrikautisista kerrostumista alkaen. Yleisiä fossiileja on suvuissa Actinomma (kuvia), Heliosphaera ja Hexadoridium (kuvia).
Varhaiset sädejalkaiset ovat selvästi erilaisia kuin myöhemmät; niillä on erilainen piitukirangan rakenne ja siinä tuskin on piikkimäisiä ulokkeita. 90 % kaikista tunnetuista säde-eliölajeista on sukupuuttoon kuolleita, ja niitä käytetään geologiseen ajanmääritykseen, öljynetsinnässä ja päätelmien tekoon muinaisista ilmasto-oloista. Merien happamoituminen eli hiilidioksidin liukeneminen meriveteen hajottaa piirangat, mikä on merkki lisääntyneestä tulivuoritoiminnasta. Nykyään ihmistoiminnan lisäämä hiilidioksidin vaikutus näkyy tarkasteltaessa säde-eliöitä mikroskoopilla.
Sädejalkaisten pääjakson taksonomia sisältää seuraavat kehityslinjat eli kladit:
Luokka 1. Polycystinea. Piipitoisen tukirangan muodostavia. Valtaosa fossiilisista lajeista kuuluu tähän ryhmään.
Lahko Nassellaria. Tukirangat kartiomaisia. Yhdeksän heimoa. – kuvia
Lahko Spumellaria. 46 heimoa, mm. Catenopylidae, Cavaspongiidae, Emiluviidae, Comberellidae, Patruliidae, Relindellidae, Spongodiscidae, Stylosphaeridae, Tholoniidae, Tritrabidae. – kuvia
Luokka 2. Spasmaria.
Lahko Acantharea. Strontiumsulfaatti-pitoinen tukiranka, joka ei fossiloidu.
Lahko Taxopodida. Vain suku Sticholonche ja siinäkin vain yksi laji S. zanclea; sisäinen tukiranka puuttuu, sen sijaan piikkejä on; eliön koko n. 200 mikronia (0,2 mm); merilaji 99 – 510 m:n syvyydessä; kaksikylkissymmetrinen, litteähkö, etupäästään leveä; aksopodit sijaitsevat riveittäin, niistä osa on liikkumattomia, suurempi osa liikkuvia kelluntavälineitä. – kuvia
Alveolata-ryhmän pääjakso Ciliophora käsitettiin aiemmin eläinkuntaan kuuluvaksi, ja sen edustajia on vanhastaan kutsuttu ripsieläimiksi. Varsin eläinmäisiä ne ovatkin liikuntakykyisinä ja pitkälle edistyneine solurakenteineen, niiden soluruumiissa on eläinten ruoansulatusjärjestelmää muistuttava osa solusuineen ja hylkyaukkoineen. Kuitenkaan ne eivät ole eläimille sukua molekyylianalyysien valossa, vaan ovat sukua alveolaattien panssarileville (Dinophyta) ja itiöeliöille (Apicomplexa) ja edustavat siten aivan eri kehityslinjaa kuin eläimet.
Tohvelieliön (Paramecium) rakenne: 1. sykkivä rakko, 2.ravintorakko, 3. isotuma, 4. pikkutuma, 5. ravintorakko tyhjenee, 6. solunielu, 7. solusuu, 8. ripset. – Franciscosp2 [Public domain].Ciliophora-pääjakso on sekin laaja ja monimuotoinen ryhmä, ja koska se ei kuulu eläinkuntaan, nimitän sitä ripsieliöiksi. Sekä tieteellinen että suomalainen nimi viittaavat solun pintaan kiinnittyviin värekarvoihin, joiden avulla eliöt liikkuvat ja pyydystävät ravintonsa. Ripsieliöt ovat petoja, jotka syövät bakteereja ja toisia aitotumaisia yksisoluisia. Värekarvat kuljettavat saaliin kohti uurretta ja sen pohjalla olevaa solusuuta eli sytostomia.
Värekarvat voivat peittää koko eliön tai voivat sijaita muutamassa rivissä tai ryhmittäin tupsumaisesti. Eliö liikkuu tiheästi sijaitsevien ripsien koordinoidun liikkeen avulla, eräillä lajeilla ripset taas voivat muodostaa jalkamaisia rakenteita sitoutumalla yhteen.
Omalaatuisin piirre ripsieliöillä on niiden kaksi tumaa: pieni mikronukleus eli pikkutuma ja iso makronukleus eli isotuma. Yhdessä solussa on kumpaakin yksi tai useampia. Konjugaatio-niminen ilmiö saa aikaan geneettistä muuntelua. Siinä kaksi ripsieliötä asettuvat rinnakkain ja osittain yhtyvät. Pikkutumassa on alunperin kaksiannoksinen kromosomisto, eli se on diploidi. Toisiinsa liittyneissä ripsieliöissä pikkutumat jakautuvat meioottisesti eli tuottavat kumpikin neljä haploidia eli kromosomistoltaan yksiannoksisista pikkutumaa. Kolme niistä kummassakin solussa katoaa, ja jäljelle jäänyt haploidi mikronukleus jakautuu kahdeksi. Ripsieliöt vaihtavat toisen pikkutumansa toisen yksilön vastaavaan ja eroavat toisistaan. Kumpikin on saanut mikrotuman genomiin uuden puoliskon toiselta, haploidit mikrotumat yhdistyvät ja muodostuu uudenlainen diploidi pikkutuma kummallekin.
Jotta tilanne ei jäisi liian yksinkertaiseksi, tapahtuu myös suvuttomassa lisääntymisessä outoja asioita. Ripsieliöt lisääntyvät jakautumalla kahtia, ja valmistautuminen siihen vaatii taas muutoksia tumajärjestelmässä. Pikkutumat alkavat jälleen jakautua mitoottisesti, ja tekevät sen peräti kolme kertaa, jolloin tulokseksi saadaan kahdeksan haploidia mikronukleusta. Alkuperäinen isotuma hajoaa ja lakkaa olemasta, ja neljä pikkutumaa muuttuu isotumiksi, joten jakautumisvalmiissa ripsieliössä on lopulta neljä isotumaa ja neljä pikkutumaa. Siinä vaiheessa eliö jakautuu kahteen kertaan, jolloin syntyy neljä uutta yksilöä, joilla kullakin on yksi iso- ja yksi pikkutuma.
Kussakin ripsieliön isotumassa on genomin moninkertainen sisältö. Isotuman geenit kontrolloivat solun arkisia toimintoja, kuten ravinnonottoa, kuona-aineiden poistoa ja vesitaloutta.
Diversiteettiä ja systematiikkaa
Ciliophora-pääjakso jaetaan kahteen alajaksoon: Postciliodesmatophora ja Intramacronucleata. Ne puolestaan jaetaan yhteensä 14 luokkaan.
Alajakso postciliodesmatophora
Heterotrichea. Luokan tieteellinen nimi tulee kreikankielen sanoista ja voitaisiin kääntää erilaisripsisiksi, mikä viittaa siihen, että suualueen värekarvat ovat erilaisia kuin muut, liikkumisesta vastaavat. Monet lajit pystyvät vetäytymään kokoon häiriintyessään. Luokkaan kuuluu eräitä kookkaimpia ripsieliöitä, kuten torvimainen Stentor (kuvia) ja Spirostomum (kuvia), sekä monia kirkkaanvärisiä muotoja, kuten punaiset Blepharisma-lajit (kuvia); Stentor coeruleus (kuvia) on sininen, ja sinipigmenttiset Eufolliculina-lajit (kuvia) muodostavat laajoja mattoja syvänmeren pohjaan. Erilaisripsiset ovat sikäli tyypillisiä alajaksonsa edustajia, että niiden soluruumiin värekarvojen liikkeitä aiheuttavat kinetosomit ovat kaksittain, ja joko etumainen tai molemmat niistä kiinnittyvät värekarvaan, ja kinetosomiparit liittyvät mikrotubulusten muodostamiin säikeisiin. Sama kinetosomijärjestelmä on myös luokassa Karyorelictea. Isotuma jakautuu ulkoisten mikrotubulusten avulla, kun yleensä ripsieliöillä se jakautuu sisäisten mikrotubulusten avulla. Video värekarvojen liikkeestä.
Karyorelictea. Luokka koostuu enimmäkseen merien interstitiaalieliöstöön kuuluvista eli hiekanjyvien välissä elävistä lajeista. Poikkeuksena on suku Loxodes (kuvia), joka elää suolattomassa vedessä. Interstitiaaliset lajit ovat suhteellisen isoja, jopa yli mm:n pituisia, ja ovat matomaisia muodoltaan. Suurin osa lajeista käyttää ravinnokseen bakteereja tai leviä ja viihtyy vähähappisissa oloissa. Kentrophoros-suvussa (kuvia) esiintyy symbioottisia rikkioksidibakteereja, joiden varassa eliön ravinto on.
alajakso intramacronucleata
Alajakson tieteellinen nimi viittaa makronukleuksen sisään muodostuvaan tumasukkulaan eli mikrotubulusrakenteeseen, joka tuman jakautuessa erottaa vastinkromosomit toisistaan ja vetää ne omiin puoliskoihinsa. Alajakson muodostavat seuraavat luokat:
Armophorea. Hapettomissa olsuhteissa eläviä ripsieliöitä, joissa on symbiontteina metanogeenisiä alkeistumaisia. Monet lajit ovat vapaina eläviä (esim. Brachonella, kuvia; Caenomorpha, kuvia; Metopus, kuvia), toiset taas eläinten, etenkin hyönteisten ja sammakkoeläinten, ruoansulatuselimistössä eläviä (esim. Nyctotherus, kuvia).
Litostomatea. Luokkaan kuuluu petoja (esim. Didinium, kuvia), selkärankaisten ruoansulatuselimistössä eläviä endosymbiontteja (esim. Balantidium coli, ainoa ihmisessä loisiva ripsieliö; kuvia) sekä vapaana eläviä lajeja. Liikuntayksikkö eli kinetidi koostuu yhdestä värekarvasta ja siihen liittyvistä rakenteista, joilla on luokalle ominainen hienorakenne. Sytostomi sijaitsee solun kärjessä.
Colpodea. Noin 200 suolattoman veden ja maaperän lajia, joiden värekarvat ovat tavallisesti kaikki samanlaisia. Sytostomi sijaitsee solun kärjessä tai alapinnalla. Monet lajit ovat epäsymmetrisiä. Suvun Colpoda (kuvia) lajit ovat munuaisen muotoisia ja elävät runsasravinteisissa ympäristöissä.
Nassophorea. Vapaana enimmäkseen makeassa vedessä, mutta myös merissä ja maaperässä eläviä. Sytostomi sijaitsee alapinnan etupäässä. Kuvia.
Phyllopharyngea. Tavallisesti värekarvaisia vain alapinnaltaan, osa lajeista kiinni-istuvia eli sessiilejä ja aikuisvaiheessaan erikoisen muotoisia (esim. Suctoria; kuvia). Luokan nimi johtuu mikrotubuluksista muodostuneista nauhoista, jotka ympäröivät solusuuta (niistä käytetään nimitystä phyllae). Enimmäkseen luokan lajit ovat vapaana eläviä, mutta on myös loisia esim. simpukoissa.
Prostomatea. Vain kaksi sukua: Coleps (kuvia) ja Pelagothrix (kuvia).
Plagiopylea. Luokka on vähälajinen ja sisältää muutamia hapettomissa oloissa eläviä eli anaerobisia lajeja. Värekarvoitus on tiheä. Kuvia
Oligohymenophorea. Runsaslajinen luokka, jonka lajeilla on tavallisesti alapinnalla uurre, jossa sytostomi sijaitsee solun muita osia peittävistä eroavine värekarvoineen. Solua peittää usein tiheä värekarvoitus. Lajit ovat laajalle levinneitä, ja niitä tavataan enimmäkseen suolattomissa vesissä saalistaen pienempiä eliöitä. Esim. tohvelieliöt (Paramecium, kuvia) kuuluvat tähän luokkaan.
Protocruziea. Luokassa on yksi kuusilajinen suku: Protocruzia (kuvia).
Spirotrichea. Luokka on runsaslajinen ja omaleimainen. Suuvärekarvat ovat silmiinpistäviä, ja soluruumiin värekarvat ovat usein kasvaneet yhteen sukasiksi (kirruksiksi) tai niitä on vähän tai ne jopa puuttuvat. Sukaselliset lajit ovat yleisiä kaikkialla maaperässä, makeassa ja suolaisessa vedessä. Ne ovat tav. litteitä ja ryömivät, uivat ja saalistavat vatsapuolen sukasilla; selkäpuolelta ne puuttuvat. Sukasettomat ja värekarvattomat lajit ovat yleensä pienikokoisempia ja enimmäkseen merissä eläviä. Niihin kuuluu heimo Tintinnidae (kuvia), jotka ovat kuorellisia ja siten hyvin säilyneet fossiileina.
Cariacotrichea. Vain yksi laji: Cariacothrix caudata (kuvia).
Mesodiniea. Systemaattiselta asemaltaan epävarma ryhmä, mahdollisesti oma alajaksonsa. Kuvia.
Malaria on menneinä aikoina ollut tuberkuloosin ohella tappavin infektiotauti. Malaria väheni 1960-luvulla malariahyttysiä (Anopheles; kuvia) tappavien hyönteismyrkkyjen ja malarialoisia (Plasmodium) tappavien lääkkeiden avulla. Sittemmin syntyi sekä hyttysten että loisen vastustuskykyisiä muotoja, ja malaria on jälleen lisääntynyt. Noin 200 miljoonaa ihmistä on saanut tartunnan tropiikissa, ja 600 000 kuolee malariaan joka vuosi.
Malarian vastaisen rokotteen kehittämistä vaikeuttaa se, että loinen elää pääasiassa punaisten verisolujen sisällä ja on immuunijärjestelmän saavuttamattomissa. Lisäksi malarialoinen unitautiloisen (Trypanosoma, ks. ekskavaatit) tapaan vaihtelee jatkuvasti pintaproteiinejaan. Kuitenkin vuonna 2015 Euroopassa hyväksyttiin maailman ensimmäinen malariarokote, mutta se antaa vain osittaisen vastustuskyvyn malariaa vastaan. Tämä rokote näet kohdistuu niihin muunteleviin pintaproteiineihin. Nykyään yritetään kehittää rokotteita, jotka kohdistuisivat muihin mahdollisiin loissolun kohteisiin.
Malarialoisen elinkierto. – Plasmodium_zyklus.png: The original uploader was Chb at German Wikipedia.Plasmodium_lifecycle_PHIL_3405_lores.jpg: * Photo Credit:Content Providers(s): CDC/Alexander J. da Silva, PhD/Melanie Moserderivative work: RicHard-59 [Public domain]Yllä oleva kuva antaa käsityksen malarialoisen monimutkaisesta elinkierrosta, joka hieman yksinkertaistaen kulkee seuraavasti: Infektoitunut malariasääski pistää ihmistä ja injisoi loisen sporotsoiitteja ihmiseen. Sporotsoiitit kulkeutuvat maksan soluihin, joissa ne jakautuvat ja muodostavat merotsoiitteja, jotka tunkeutuvat punaisiin verisoluihin. Merotsoiitit jakaantuvat punaisissa verisoluissa, joista ne purkautuvat suurin joukoin tietyin ajanjaksoin ja aiheuttavat horkan ja kuumeen. Eräät merotsoiitit muodostavat gametosyyttejä, muut tunkeutuvat uusiin punasoluihin. Kun malariasääski imee verta tartunnan saaneesta ihmisestä, se saa loisen gametosyyttejä elimistöönsä. Siellä gametosyyteistä muodostuu sukusoluja eli gameetteja. Sääsken suolessa tapahtuu hedelmöitys ja tsygootista kehittyy ookysti, josta vapautuu tuhansia sporotsoiitteja, jotka vaeltavat sääsken sylkirauhaseen, mistä ne jälleen joutuvat ihmiseen jne …
Malarialoinen (Plasmodium) on esimerkki Apicomplexa-pääjakson eliöistä.
Apicomplexa
Apicomplexa on suuri pääjakso, joka sisältää loisivia alveolaatteja. Suurimmalla osalla sen lajeista on ainutlaatuinen soluelin, joka koostuu eräänlaisesta plastidista (apikoplasti) ja solun kärjessä olevasta monimutkaisesta rakenteesta. Soluelin on sopeutuma, jonka avulla parasiitti tunkeutuu isäntäsoluun. Apicomplexa-taksonin nimi on johdettu kahdesta latinalaisesta sanasta: apex ’kärki, nokka’ ja complexus ’ympäröiminen, syleily’. Se viittaa sporotsoiitin edellä mainittuun soluelinapparaattiin.
Apicomplexa-eliöt ovat yksisoluisia ja itiöitä muodostavia. Ennen, kun oli olemassa alkueläinten pääjakso (Protozoa) eläinkunnassa (Animalia), niitä sanottiin itiöeläimiksi (Sporozoa). Ehkä niitä nyt voisi kutsua itiöeliöiksi, kun eivät kuulukaan eläimiin. Liikuntaelimiä, kuten siimoja tai valejalkoja, esiintyy vain tietyillä sukusoluilla. Kuitenkin enin osa itiöeliöistä on liikkuvia eräänlaisen liukumismekanismin avulla.
Kaikki itiöeliöt ovat eläinten sisäloisia. Ne aiheittavat eläimissä monia sairauksia kuten babesioosia (Babesia; kuvia), malariaa (Plasmodium; kuvia), kryptosporidioosia (Cryptosporidium parvum; kuvia), kyklosporiaasia (Cyclospora cayetanensis; kuvia), isosporiaasia (Isospora belli; kuvia) ja toksoplasmoosia (Toxoplasma gondii; kuvia).
Kaikilla Apicomplexa-pääjakson lajeilla on infektion aiheuttava elämänvaihe, jota sanotaan sporotsoiitiksi, ja sillä on kärjessään apikaalikompleksi- niminen rakenne, josta jo edellä mainittiin. Apikaalikompleksi käsittää sarjan spiraalimaisesti järjestyneitä mikrotubuluksia, erittävän kappaleen ja yhden tai useampia rengasmaisia muodostumia, joiden sisäpuolella on toisinaan ohuita elektronipitoisia erityselimiä. Kaikkialla solussa on pallomaisia soluelimiä, joiden eritteet vapautuvat isäntäsoluun loisen tunkeuduttua siihen.
Itiöeliöiden tuma on haploidi eli kromosomistoltaan yksiannoksinen. Vaihteleva määrä siimoja on vain liikkuvissa gameeteissa, ja ne suuntautuvat solun takaosaa kohti. Niiden tyvessä on tyvijyvänen eli kinetosomi, joka organisoi siimojen liikkeet. Mitokondrioiden kristat ovat putkimaisia. Solussa on myös Golgin laite. Joillakin lajeilla on värittömiä plastideja. Solukalvo koostuu kolmesta kerroksesta, joita pienet huokoset lävistävät.
Lisääntyminen tapahtuu tavallisesti jakautumalla mitoottisesti. Itiöeliöiden mitoosissa ainutlaatuista on se, että tumasukkula, joka vetää kromosomit erilleen tytärsoluihin, on tavallisimmin tumansisäinen, kun se tavallisesti eukaryooteilla muodostuu tuman ulkopuolelle. Meioosi, jonka tuloksena syntyy haploideja sporotsoiitteja, tapahtuu vasta hedelmöityneessä munasolussa eli tsygootissa.
Useimmilla Apicomplexa-pääjakson lajeilla on monimutkainen elämänkierto, jossa on sekä suvullista että suvutonta lisääntymistä. Tyypillisesti isäntäsoluun tunkeutuu sporotsoiitteja, jotka saavat isännän lopulta hajoamaan, jolloin merotsoiitit vapautuvat infektoimaan uusia soluja. Tämä voi toistua useita kertoja ennen kuin syntyy gamontteja, jotka tuottavat sukusoluja. Sukusolut yhtyvät ja muodostavat kystejä, lepomuotoja, jotka voivat levitä uuteen isäntäeläimeen. Tästä perusmallista on monenlaisia muunnelmia, ja monilla itiöeliöillä on enemmän kuin yksi isäntä.
luokittelu
Apicomplexa-pääjaksoon kuuluu kolme pääryhmää, joita voidaan pitää luokkina:
Suippoloisiot (Gregarisina; kuvia): tavallisesti nivelmatojen, niveljalkaisten ja nilviäisten loisia; usein suolistoloisia; tyypillisessä elinkierrossa trofotsoiitti kehittyy isäntäsolussa skitsontiksi, joka jakautuu merotsoiiteiksi; isäntäsolun hajotessa ne tunkeutuvat toisiin soluihin; tämä toistuu, kunnes sukusolut syntyvät; ne vapautuvat sen isäntäsolun hajotessa, jossa ovat muodostuneet; gameetit muodostavat kystejä, jotka voivat siirtyä uusiin isäntäeläimiin.
Kokkidiot (Coccidia; kuvia): yleensä selkärankaisten loisia; sekä suolen epiteelisoluissa että muiden kudosten soluissa; elinkierto kolmivaiheinen: merogonia, gametogonia ja sporogonia; gametogonia ainutlaatuinen, siinä trofotsoiiteista osa suurenee makrogameeteiksi, osa jakautuu liikkuviksi mikrogameeteiksi; tsygootti muuttuu kystiksi, joka vapautuu isäntäeläimestä; elinkierto haplontti, vain tsygootti lyhytaikainen diploidi vaihe.
Aitotumaiset eli eukaryootit jakautuvat neljän pääryhmän kesken, joista käsillä olevassa SAR-ryhmässä on ainakin kolme alaryhmää: sukasiimaiset (Stramenopila), alveolaatit (Alveolata) ja valejalkaiset (Rhizaria). Panssarilevät (Dinophyta) ovat alveolaatteja.
Alveolaatit ovat yksisoluisia, ja niiden ominaisuuksista huomiota herättävimpiä ovat alveolit, jotka ovat kalvon ympäröimiä säkkimäisiä muodostumia aivan solukalvon eli plasmamembraanin alla. Alveolaatit ovat runsaita monenlaisissa elinympäristöissä, ja niissä on niin omavaraisia, yhteyttäviä, kuin toisenvaraisiakin eliöitä. Rymä on hyvin monimuotoinen ja käsittää vapaana eläviä siima- ja ripsieliöitä sekä loisia.
Panssarilevien pääjakso
Panssarilevät, joita kutsutaan myös dinoflagellaateiksi, ovat usein selluloosalevyjen muodostaman ulkoisen tukirangan peittämiä. Panssarin uurteissa sijaitsevat kaksi siimaan saavat eliön liikkumaan kieppumalla.
Panssarileviä elää sekä merissä että sisävesissä, enimmäkseen kuitenkin merten planktonissa.
Panssarilevien kantamuodot ovat saattaneet syntyä sekundaarisen endosymbioosin avulla. Silti karkeasti ottaen puolet lajeista on pelkästään toisenvaraisia. Muut ovat tärkeä osa fytoplanktonia eli keijustoa, joka käsittää yhteyttävät bakteerit ja levät. Monet fotosyntetoivista panssarilevistä ovat miksotrofeja: ne voivat yhteyttämisen ohella ottaa ravintoa myös solun ulkopuolelta heterotrofien tapaan. On myös selkärangattomien merieläinten soluissa eläviä endosymbiontteja.
Panssarilevät aiheuttavat välillä huomattavia kukintoja ja värjäävät veden punaiseksi tai ruskehtavaksi. Väri johtuu karotenoideista, jotka ovat panssarilevien plastidien yleisin pigmentti. Tietyt panssarilevälajit erittävät myrkkyjä, jotka ovat aiheuttaneet massiivisia kalojen ja selkärangattomien eläinten kuolemia. Joskus ihmisetkin voivat saada jopa kuolettavia myrkytyksiä syötyään nilviäisiä, jotka ovat keränneet kudoksiinsa näitä myrkkyjä.
evoluutio ja luokittelu
Panssarilevien fossiilisia lajeja tunnetaan noin 2000, joista vanhimmat varmuudella tunnistetut ovat eläneet 410 miljoonaa vuotta sitten siluurikaudella. Prekambrikaudelta on peräisin paljon mikrofossiileja, joista osa saattaa kuulua panssarilevien kehityslinjaan. Suurimmillaan monimuotoisuus oli ilmeisesti 110 – 80 miljoonaa vuotta sitten liitukauden jälkipuoliskolla.
Bioluminesenssia rantautuvassa aallossa. – catalano82 [CC BY 2.0 (https://creativecommons.org/licenses/by/2.0)].Nykyisiäkin panssarilevälajeja tunnetaan noin 2000, hieman yli. Dinophyta-pääjaksossa on viisi luokkaa ja 20 lahkoa (Ruggiero ja muut 2015: A Higher Level Classification of All Living Organisms. PLoS ONE 10:4). Tunnetuimpia sukuja ovat mm. Ceratium (kuvia), Peridinium (kuvia), Gymnodinium (kuvia), Symbiodinium (kuvia) ja Noctiluca (kuvia). Viimeksi mainittu on tunnettu kyvystään tuottaa valoa kemiallisesti; ilmiötä kutsutaan bioluminesenssiksi.
Polykrikoskofoidii. – English: Light micrograph of an extruded nematocyst of Polykrikos kofoidii. Scale bar = 10 microns. This image is lightly edited to remove distracting panel label from Panel L of Figure 1 from Hoppenrath et al. 2009, Molecular phylogeny of ocelloid-bearing dinoflagellates (Warnowiaceae) as inferred from SSU and LSU rDNA sequences, BMC Evolutionary Biology 9:116. doi:10.1186/1471-2148-9-116. This file is licensed under the Creative Commons Attribution 2.0 Generic license.
Seuraavassa ote Niko Kettusen kirjoituksesta Viikon eläin, Helsingin sanomat 30.4.2018 (yllä olevassa kuvassa kirjoituksessa mainittu panssarilevä).
PIENELIÖISTÄ löytyisi aivan hurjia lätkämaskotteja. On esimerkiksi olemassa hermomyrkkyä tuottava, panssaroitu otus, joka saalistaa uhrejaan harppuunalla.
Se on hallinnut maailman meriä miljoonia vuosia ja pystyy yöllä valaisemaan kokonaisia alueita hohtamalla valoa. Se on niin ovela, ettei siitä oikein tiedä, onko se eläin vai kasvi. Ja kaikki tämä yhdessä ainoassa solussa!
Kyse on tietysti panssarilevistä eli dinoflagellaateista. Niitä tunnetaan tuhansia lajeja, ja kaikki ovat aivan mahtavia. Esimerkiksi Polykrikos kofoidii -panssarilevä viettää päivänsä ampumalla muita leviä ja eliöitä pienellä harppuunalla, joka on kerällä otuksen sisällä.
Harppuuna lamauttaa uhrin, minkä jälkeen Polykrikos hinaa sen syötäväksi. Se saalistaa myös niitä panssarilevälajeja, jotka tuottavat harmillisia leväkukintoja.
SAALISTAVIEN panssarilevien esivanhemmat saivat ravintoa rauhanomaisesti yhteyttämällä. Sitten ne piirsivät evoluutiossa pelikuviot uusiksi ja rupesivat tappamaan. Joillekin panssarileville on kehittynyt todellinen yksisoluinen konekivääri – niillä voi olla jopa 15 harppuunaa.
Yksilönä panssarilevä on tietysti aika vähäinen, mutta yksilösuorituksista muodostuu voimakas joukkue. Maailman merissä raivoaa geologisten ajanjaksojen yli ulottuva leväkukintojen kuolinkamppailu, jossa mikrobien mongoliarmeijat ottavat yhteen. Mikä dynastia, mikä taustatarina!
Merenrantojemme näkyvin levä viherlevävyöhykkeiden ohella on rakkolevä (Fucusvesiculosus, kuvia). Ruskeana se vaikuttaa kuolleelta kasvilta, mutta ulkonäkö pettää. Rakkolevä kuuluu ruskolevien pääjaksoon (Phaeophyta), jonka lajeista suurin osa on ruskeita tai kellanruskeita, sillä runsaat karotenoidit ja ruskea fukoksantiini peittävät yhteyttämispigmentin, viherhiukkasten lehtivihreän. Rakkolevän lisäksi niukkasuolaisessa merivedessämme elää kolmisenkymmentä ruskolevälajia.
Rakkolevä on Itämeren tärkein levä – se on ainoa suuria, yhtenäisiä ja monivuotisia kasvustoja muodostava levä. Se puuttuu vähäsuolaisimmista osista Suomenlahden perukasta ja Perämerestä sekä jokisuistoista. Parhaimmillaan se voi lounaisilla merialueillamme saavuttaa jopa metrin pituuden, mutta on tavallisesti paljon pienempi. Parilliset kaasurakkulat sekovarren haarojen päissä pitävät rakkolevän pystyasennossa; pienimmiltä yksilöiltä ne puuttuvat. Kaasurakkuloiden ohella sekovarren kärjissä on muna- ja siittiöpesäkkeistöjä, joista vapautuu siimallisia, itsenäisesti uivia siittiöitä tai virtausten mukana ajelehtivia munasoluja. Jos hedelmöitynyt munasolu päätyy kiinteälle alustalle, se kiinnittyy paikalleen ja kasvaa uudeksi yksilöksi.
Rakkolevän tiheät kasvustot ulottuvat kiinteäpohjaisilla kasvupaikoilla 7 – 8 m:n syvyyteen. Irronneina niitä voi kertyä salmien ja lahtien liejupohjillekin paksuiksi lautoiksi, joiden pintaosa elää ja kasvaa, vaikka alaosat lahoavat.
Itämeren rehevöityminen on vähentänyt rakkoleväkasvustoja. 1970-luvun lopulla ne katosivat Turun seudun ja Ahvenanmaan itäpuolisesta saaristosta, mutta palasivat muutaman vuoden kuluttua. Tämäkin ilmiö aiheutui ihmistoiminnan, erityisesti maanviljelyksen, aiheuttamasta rehevöitymisestä. Nopeakasvuiset rihmalevät runsastuivat ravinnelisäyksen vuoksi ja niiden mukana leväsiira, joka syö nuorena rihmaleviä ja vanhemmiten rakkolevää. Rihmalevät peittivät rakkolevän ja rusastunut siirakanta söi kasvustot. Aikuisilla leväsiiroilla ei ollut lopulta rakkolevän antamaa suojaa, jolloin kalat söivät niitä runsain mitoin. Kun siirakanta väheni, palasi rakkolevä.
Rakkoleväkato vaikutti myös kalakantoihin, jotka romahtivat, kun ne olivat menettäneet rakkolevän suoman lisääntymisturvan. Tämä näkyi erityisesti ahvenen ja hauen määrissä. Rakkolevä suo myös monille muille eliöille sopivan elinympäristön, kuten monille kalalajeille, leväkatkaravulle ja muille äyriäisille, sinisimpukoille, lima- ja leväkotiloille, levärupena tunnetuille sammaleläimille, merirokolle ja lovéninrunkopolyypille.
Jouhilevä (Chorda filum, kuvia) on toinen rannikoittemme huomiota herättävä ruskolevä. Sen sekovarret ovat pitkiä ja kengännauhamaisia.
Ruskolevien pääjakso
Ruskolevien pääjaksoon (Phaeophyta; kuvia) kuuluvat eliöt ovat kaikki monisoluisia, ja lähes kaikki suolaisen veden eläjiä. Koko vaihtelee kovasti: on mikroskooppisen pieniä ja rihmamaisia sekä kymmenien metrien pituisia ja sekovarreltaan erilaistuneita ja kaikkea niiden väliltä. Useimmat elävät viileiden ja kylmien merien rannikkovesissä pohjaan kiinnittyneinä.
plastidit
Kuten piilevillä (Bacillariophyta) on ruskolevienkin plastideissa neljä kalvoa, ja tylakoidit sijaitsevat kolmen ryhmissä. Kussakin solussa voi olla yksi tai useampia plastideja ja niissä yksi tai useampia pyrenoideja eli proteiinijyväsiä, joiden ympärillä on usein tärkkelysjyväsiä. Tylakoidien pigmentit ovat klorofyllit a ja c sekä erilaiset karotenoidit, joista ennen kaikkea fukoksantiini aiheuttaa ruskoleväsolun värin. Yhteyttämistuotteet varastoituvat plastidin ulkopuolelle krysolaminariinina.
solut
Ruskoleväsolua ympäröi solunseinä, jonka sisäkerros on pääasiassa selluloosaa ja ulkokerros hyytelömäisiä polysakkarideja (sokereita) ja alginiinihappoja sekä niiden suoloja (alginaatteja). Kasvulliset solut ovat siimattomia. Sen sijaan parveiluitiöissä ja gameeteissa eli sukusoluissa on yleensä kaksi solun kylkeen kiinnittyvää siimaa. Kulkusuuntaan osoittavassa siimassa on sukasia eli mastigoneemoja, kuten yleensä sukasiimaisten ryhmässä (Stramenopila), johon ruskolevät kuuluvat mm. pii- ja kultalevien kanssa. Toinen siima on sileä ja osoittaa päinvastaiseen suuntaan. Sen tyvellä voi olla pullistuma, joka muodostaa valonaistimen plastidissa olevan silmätäplän kanssa.
sekovarsi
Joillakin ruskolevälajeilla on pieni, rihmamainen sekovarsi, jonka kaikki solut kykenevät jakautumaan. Monilla on litteitä, alustaa myöten kasvavia tai tupsumaisia sekovarsia. Suurikokoisten lajien sekovarret ovat usein erilaistuneet kasvien juuria, vartta ja lehtiä muistuttaviksi osiksi. Sekovarressa ei ole erilaistuneita solukoita eikä siis tukisolukoitakaan, ja siksi ison levän pystyasennon takaamiseksi siinä on usein yksi tai useampia kaasurakkuloita, pneumatokystejä. Rihmamaisten kasvu on yleensä hajanaista tai tapahtuu rihman tyvellä. Isokokoisilla on usein välikasvuvyöhykkeitä, ja rakkolevät kasvavat pääasiassa kärjistään. Monilla isoilla ruskolevillä on ulkopinnalla ns. meristodermi, joka on jakaantumiskykyisten solujen kerros.
elämänkierto ja lisääntyminen
Ruskolevien elinkierto on usein varsin monimutkainen. Yksinkertaisimmasta päästä on diplohaplonttinen eli diplobionttinen elämänkierrosta, jolla tarkoitetaan, että meioosi eli kromosomiston puolittuminen tapahtuu sukupuolisolujen muodostuessa eikä itiöiden muodostuessa. Niin ollen levän kaikki elämänvaiheet sukusoluja lukuun ottamatta ovat diploideja eli kromosomisto esiintyy kaksinkertaisena. Tämä tilanne on mm. rakkolevillä.
Monimutkaisempi elinkierto on sukupolvenvuorottelu, jossa on monisoluiset haploidi ja diploidi sukupolvi. Kun haploidi vaihe on yksisoluinen (sukusolut eli gameetit), ei puhuta sukupolvenvuorottelusta (itse asiassa ihminen on rakkolevän tapaan tällainen). Sukupolvenvuorottelussa esim. Laminaria-lajeilla diploidia vaihetta nimitetään sporofyytiksi, koska se tuottaa itiöitä eli spooreja. Ne ovat haploideja eli meiospooreja, meioosiksi nimetyn solujen jakaantumisen tuottamia itiöitä (meioosi tuottaa kromosomistoltaan yksiannoksisia soluja, mitoosi puolestaan kaksiannoksisia). Lisäksi nämä meiospoorit ovat siimallisia ja liikkuvia, ja siksi niitä nimitetään tsoospooreiksi. Ne kehittyvät edelleen haploidisiksi gametofyyteiksi, jotka tuottavat sukusoluja; naarasgametofyytti tuottaa munasoluja ja koirasgametofyytti siittiösoluja. Sukusolujen yhtyminen eli hedelmöitys tuottaa diploidin (kromosomistoltaan kaksiannoksisen) tsygootin, josta syntyy uusi sporofyytti, joka aikanaan taas tuottaa meiospooreja. Mainitulla esimerkkimerkkisuvulla sporo- ja gametofyytti ovat rakenteeltaan erilaisia, heteromorfisia. Jos suvullinen eli sukusoluja tuottava sukupolvi ja suvuton sukupolvi ovat rakenteeltaan samanlaisia, vaikka eroavat kromosomimäärältään, on kyse isomorfisesta sukupolvenvuorottelusta.
Laminariahyperborea. – Franz Eugen Köhler, Köhler’s Medizinal-Pflanzen [Public domain].
Itiöt syntyvät itiöpesäkkeissä eli sporangioissa. Ruskolevillä niitä on kahdenlaisia: monisoluisia ja yksisoluisia. Sukusolut syntyvät munapesäkkeissä (oogonioissa) ja siittiöpesäkkeissä (anteridioissa). Sukusolujen yhtyminen voi ruskolevillä olla kolmenlaista:
isogamiassa sukusolut ovat kooltaan, muodoltaan ja käyttäytymiseltään samanlaiset,
anisogamiassa eli heterogamiassa sukusolut poikkeavat toisistaan kokonsa ja/tai liikkuvuutensa suhteen,
oogamiassa munasolut ovat liikumattomia ja huomattavasti suurempia kuin pienet ja liikkuvat siittiösolut.
Lisäksi monet ruskolevät lisääntyvät vain suvuttomasti.
ruskolevien ekologiaa
Runsaimmat ruskoleväkasvustot löytyvät puhtaista merivesistä ja kallioisilta rannikoilta. Tunnettuja ovat Pohjois-Amerikan länsirannikon sankat levämetsät, joissa kasvaa useiden kymmenien metrien mittaisiksi tulevia Macrocystis– ja Nereocystis-lajeja, jotka kiinnittyvät merenpohjaan. Pienemmät ruskolevälajit esim. Pohjois-Atlantilla kiinnittyvät usein kiviin tai kasvavat päällyslevinä (epifyytteinä) isompien pinnalla. Onpa ruskolevissä myös loisia, jotka ovat pitkälle erikoistuneita endofyyttejä muiden merilevien sekovarsissa.
Päärynälevä (Macrocystispyrifera). – This image is in the public domain because it contains materials that originally came from the U.S. National Oceanic and Atmospheric Administration, taken or made as part of an employee’s official duties.
ruskolevät ihmisen taloudessa
Joitakin ruskoleviä kerätään ja kasvatetaan ihmisravinnoksi erityisesti Kiinassa ja Japanissa. Vuosittain kerätty leväsato lähestyy kahta miljoonaa tonnia. Siitä suurin osa syödään sellaisenaan, lopuista valmistetaan alginaatteja. Niitä käytetään hyytelöimisaineina elintarviketeollisuudessa sekä esimerkiksi lääkekapselien ja maalien raaka-aineena.
monimuotoisuus ja luokittelu
Ruskolevälajeja tunnetaan noin 2000 lajia ja runsaat 250 sukua. Kaikki katsotaan kuuluviksi samaan ruskolevien luokkaan, Phaeophyceae, joka vuonna 2014 julkaistun luokituksen mukaan jaetaan neljäksi alaluokaksi ja lähes 20 lahkoksi.
DNA-sekvenssien perusteella ruskolevien luokan/pääjakson iäksi on arvioitu 200 miljoonaa vuotta, mutta varmuudella tunnistettuja fossiileja tunnetaan vain mioseenikaudelta 7 – 15 miljoonan vuoden takaa. Pehmeät sekovarret eivät juuri fossiloidu, ja ruskolevien kehityshistoriasta tiedetään vain vähän. Yhteinen kantamuoto lienee piilevien (Bacillariophyta) kanssa, ja joitakin yhteisiä piirteitä niillä on eräiden nykyään elävien, perinteisesti kultaleviin (Chrysophyta) luettujen levien kanssa.
Ruskolevien alaluokat, lahkot ja heimot:
Alaluokka 1. Discosporangiophycidae.
Lahko 1.1. Discosporangiales. Heimot Choristokarpaceae ja Discosporangiaceae. Alaluokka 2. Ishigeophycidae.
Lahko 2.1. Ishigeales. Heimot Petrodermataceae ja Ishigeaceae. Alaluokka 3. Dictypophycidae.
Lahko 3.1. Syringodermatales. Heimo Syringodermataceae.
Lahko 3.2. Onslowiales. Heimo Onslowiaceae.
Lahko 3.3. Dictyotales. Heimo Dictyotaceae.
Lahko 3.4. Sphacelariales. Heimot Lithodermataceae, Phaeostrophiaceae, Sphacelodermataceae, Stypocaulaceae, Cladostephaceae ja Sphacelariaceae. Alaluokka 4. Fucophycidae.
Lahko 4.1. Desmarestiales. Heimot Arthrocladiaceae ja Desmarestiaceae.
Lahko 4.2. Sporochnales. Heimo Sporochnaceae.
Lahko 4.3. Ascoseirales. Heimo Ascoseiraceae.
Lahko 4.4. Scythothamnales. Heimot Asteronemataceae, Bachelotiaceae ja Splachnidiaceae.
Lahko 4.5. Laminariales. Heimot Phaeosiphoniellaceae, Akkesiphycaceae, Pseudochordaceae, Chordaceae, Agaraceae, Laminariaceae, Aureophycaceae ja Alariaceae.
Lahko 4.6. Asterocladales. Heimo Asterocladaceae.
Lahko 4.7. Ectocarpales. Heimot Sorocarpaceae (asema epävarma), Adenocystaceae, Scytosiphonaceae, Petrosporangiaceae, Ectocarpaceae, Acinetosporaceae ja Chordariaceae.
Lahko 4.8. Stschapoviales. Heimot Stschapoviaceae, Halosiphonaceae ja Platysifonaceae.
Lahko 4.9. Tilopteridales. Heimot Masonophycaceae (asema epävarma), Tilopteridaceae.
Lahko 4.10. Ralfsiales. Heimot Mesosporaceae, Neoralfsiaceae ja Ralfsiaceae.
Lahko 4.11. Nemodermatales. Heimo Nemodermataceae.
Lahko 4.12. Fucales. Heimot Bifurcariopsidaceae, Hormosiaceae, Notheiaceae, Seirococcaceae, Xiphophoraceae, Sargassaceae, Durvillaeaceae, Himanthaliaceae ja Fucaceae.
Kuvia ja kuvauksia Suomen ruskolevistä löytyy tästä.
Kultalevien pääjakso, Chrysophyta, kuuluu sukasiimaisiin (Stramenopila), joiden merkittävä tuntomerkki on solun kaksisiimaisuus, ja se, että toinen siima on sukaspintainen, ns. mastigoneemojen peittämä. Kultalevien lisäksi sukasiimaisia ovat mm. piilevät ja ruskolevät. Kultaleviin kuuluu noin 1000 lajia, jotka ryhmitellään noin 200 suvuksi.
Kultalevien väritys johtuu keltaisista ja ruskeista karotenoideista. Kultalevät ovat useimmiten yksisoluisia. Yleensä solun toisessa päässä on kaksi siimaa (esim. Ochromonas; kuvia), mutta tunnetaan myös siimattomia eli kokkaalisia (esim. Chrysosphaera; kuvia) ja amebamaisia (esim. Rhizochrysis; kuvia) kultaleviä. Kultaleväsoluissa on yleensä silmätäplästä ja sileän uintisiiman tyvipullistumasta koostuva valonaistinelin (fotoreseptori). Solun pinnalla on usein piipitoisia suomuja.
Dinobryon divergens -kolonia. – Dr. Imre Oldal [CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0)].Kultalevissä on myös kolonioissa eläviä (esim Dinobryon suolattomassa vedessä; kuva yllä), joko uintisiimallisia (esim. Synura) tai siimattomia, rakennetasoltaan ns. kokkaalisia (esim. Chrysocapsa; kuvia). Harvat ovat rihmamaisia tai niillä yksinkertainen sekovarsi (Phaeothamnion,kuvia, ja Thallochrysis).
Synuraasmundiae. – Dima Kapustin [CC BY 4.0 (https://creativecommons.org/licenses/by/4.0)].
Monet kultalevälajit uiskentelevat suolattoman tai suolaisen veden planktonissa eli keijustossa. Useat lajit viihtyvät karuissa, happamissa ja viileissä vesissä, ja niillä onkin tärkeä merkitys maamme vesien perustuotannossa. Hyvin pienillä planktonlevillä, kuten Pelagococcus-lajeilla (kuvia), on tärkeä rooli valtamerten perustuotannossa; joillakin alueilla ne vastaavat jopa 90% perustuotannosta. Kultaleviä on myös endosymbiontteina planktisten huokoseliöiden (Foraminifera) soluissa.
Kaikki kultalevät ovat fotosynteettisiä, eräät miksotrofeja, so. ne kykenevät absorboimaan liuenneita orgaanisia yhdisteitä tai fagosytoimaan isompia ravintokappaleita, vaikka ovat myös yhteyttäviä. Jotkut lajit voivat menettää plastidinsa kasvaessaan runsasravinteisella alustalla.
Olosuhteiden huonontuessa, monet kultalevät muodostavat kestäviä kystejä, jotka voivat säilyä vuosikymmeniä. Niiden seinissä on piitä, ja muodoltaan ne ovat pyöreitä tai soikeita. Pintaa peittävät usein piikit tai muunlaiset ulokkeet. Kultaleväkystit säilyvät hyvin fossiileina, ja niitä on löydetty 80 miljoonan vuoden ikäisistä tai nuoremmista kerrostumista.
Kultalevät lisääntyvät yleensä suvuttomasti jakautumalla, eikä useimpien lajien suvullista lisääntymistä tunneta. Niillä, jotka tunnetaan, munasolut ja siittiöt ovat samankokoisia. Tätä ilmiötä nimitetään isogamiaksi, ja suvullisesti lisääntyvät kultalevät ovat siten isogamisia. Hedelmöitynyt munasolu eli tsygootti on ns. hypnotsygootti, mikä merkitsee sitä, että se tarvitsee lepovaiheen ennen kehittymistään eteenpäin.
Kultaleväsystematiikkaa
Vuonna 2014 julkaistun järjestelmän mukaan kultalevät jaetaan neljään lahkoon. Niiden ulkopuolelle jää 11 sukua, joiden asema luokittelussa on epäselvä.
Lahko Chromulinales sisältää kahdeksan heimoa. Chromulinaceae-heimossa on 11 sukua, mm. Chromulina ja Monas. – kuvia
Lahko Hibberdiales käsittää ainakin kolme sukua: Chromophyton, Hibberdia ja Lagynion. – kuvia
Keltalevät (Xanthophyta) muistuttavat rakenteeltaan ja elintavoiltaan kultaleviä. Keltaleviä tunnetaan 600 lajia ja 100 sukua. Valtaosa niistä elää suolattomassa vedessä, joitakin myös kosteassa maassa, merivedessäkin muutama. Keltalevät eivät yleensä aiheuta suuria leväkukintoja.
Myös keltalevät ovat pääosin yksisoluisia planktonleviä, yleensä siimattomia ja pallomaisia eli ns. kokkaalista rakennetasoa (esim. Chloridella, kuvia). Kolonioita muodostavat lajitkin ovat pääosin siimattomia (esim. Gloeochloris, kuvia). Vain harvat keltalevät ovat uintisiimallisia (esim. Chloromeson) tai amebamaisia (esim. Myxochloris). Pääjaksossa on myös joitakin sifonaalisia, putkimaisia lajeja (Vaucheria, Botridium) ja harvoja monisoluisia sekovarsia muodostavia (Tribonema, kuvia). Sekovarrella tarkoitetaan versomaista rakennetta, jossa ei ole erotettavissa juuria, varsia eikä lehtiä.
Kelta- ja kultalevät ovat melko samanlaisia eliöitä. Pääasiallisin ero kultaleviin nähden on solunseinän ja lepokystien hienorakenteessa ja yhteyttämispigmenteissä. Keltalevien plastidit ovat tavallisesti linssimäisiä ja fukoksantiini puuttuu (sitä on kulta- ja ruskolevillä). Luonteenomainen kellanvihreä sävy johtuu toisesta ksantiinista. Plastidien tylakoidit sisältävät karotenoideja, klorofylli a:ta ja usein vain niukasti klorofylli c:tä.
Useimmat keltalevät näyttävät lisääntyvän vain suvuttomasti jakautumalla tai suvuttomasti tuotetuilla itiöillä. Vain Vaucheria-lajien tiedetään varmuudella lisääntyvän myös suvullisesti. Niiden muna- ja siittiösolut eroavat selvästi toisistaan: munasolu on iso, siitiöt liikkuvia, kaksisiimaisia (oogamia). Tsygootin seinä paksuuntuu, ja lepovaiheen jälkeen siitä kasvaa uusi sekovarsi.
Keltalevien systematiikkaa
Keltalevät jaetaan kahteen lahkoon vuonna 2012 julkaistun systematiikan mukaan:
Lahko Tribonematales: suvut Botrydium, Bumilleriopsis, Characiopsis, Chloromeson, Heterococcus, Ophiocytium, Sphaerosorus, Tribonema, Xanthonema. – kuvia

























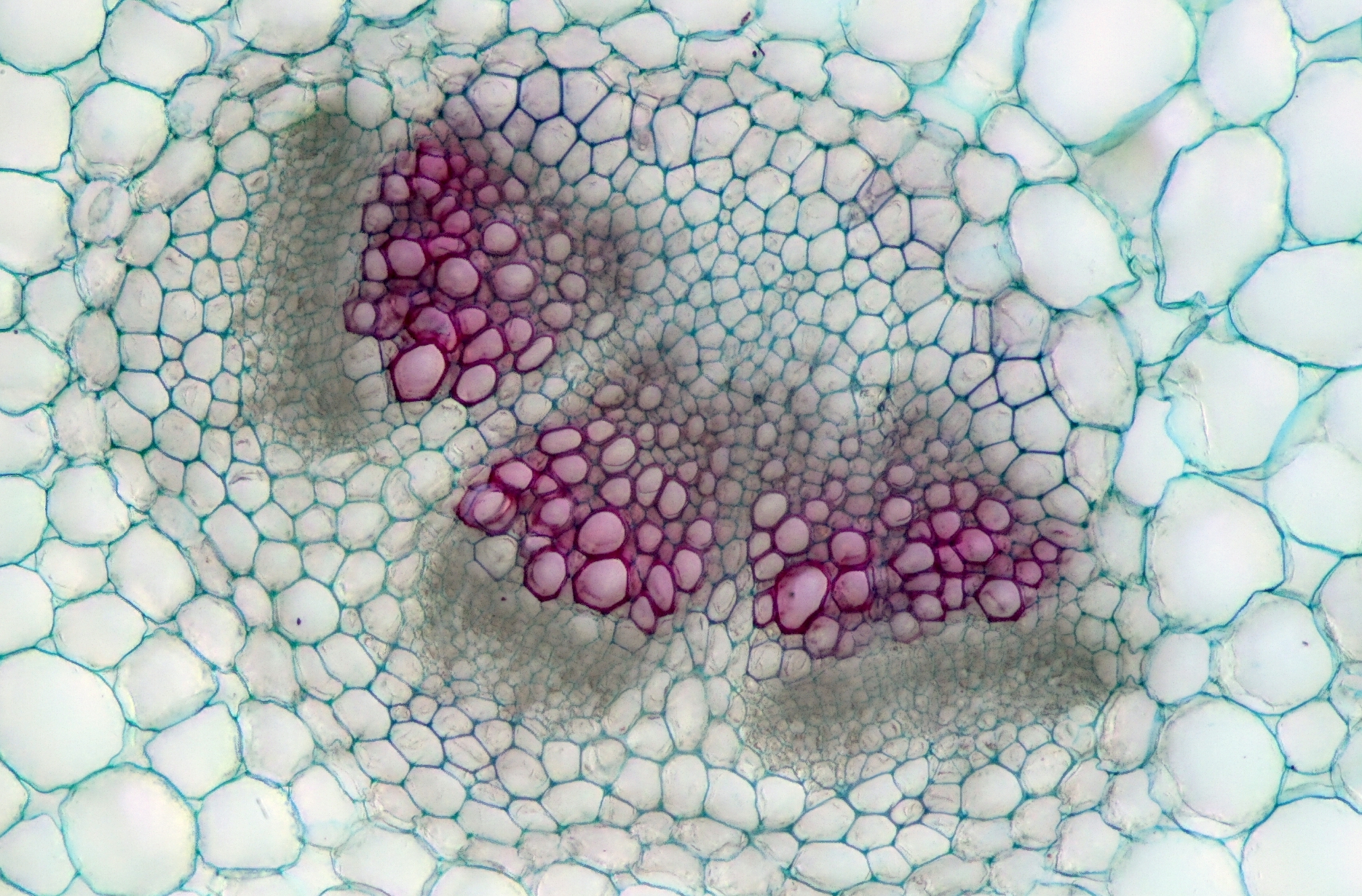





















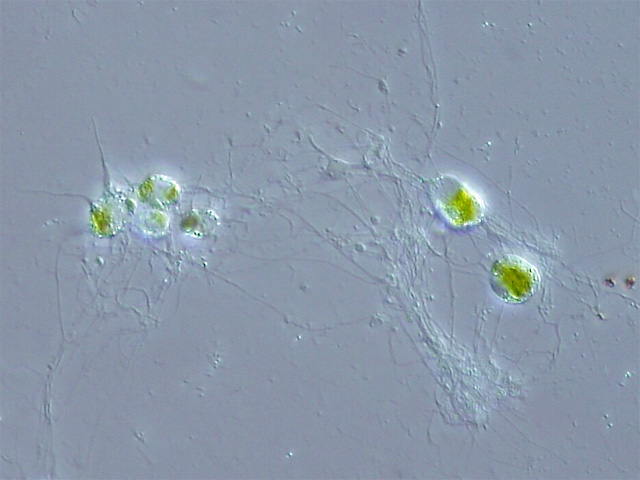









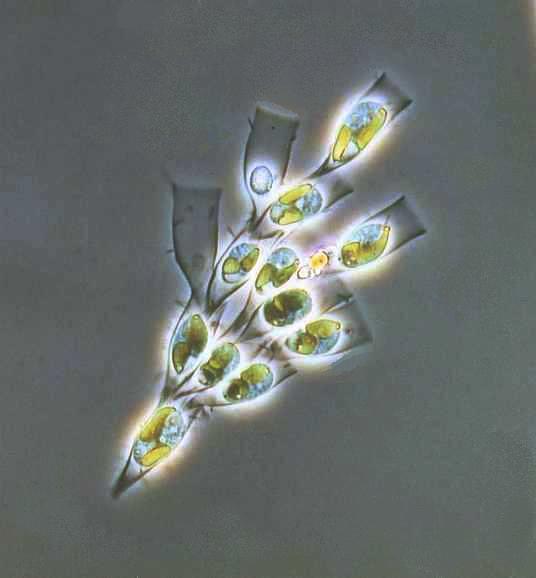

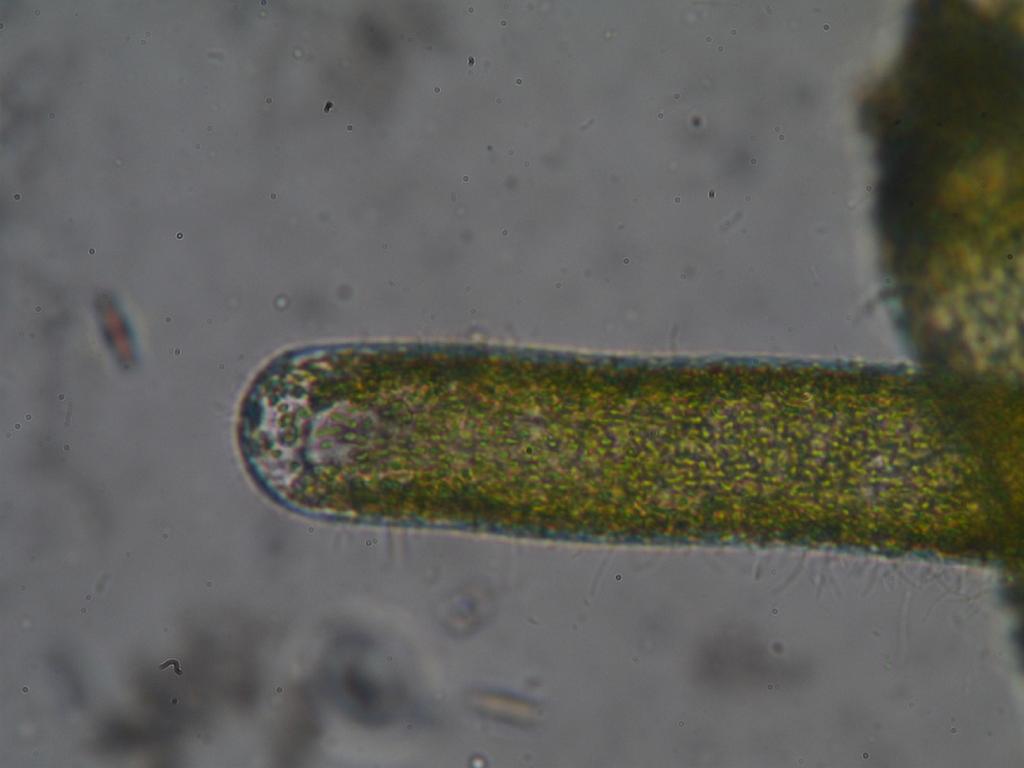
_6386.JPG){kind=link}