
Systemaattinen asema eliökunnassa:
- Esitumaiset eli tumattomat (Prokaryota)
- Aitotumaiset (Eukarya)
- Ekskavaatit (Excavata)
- SAR
- Stramenopila
- Alveolata
- Rhizaria
- Radiolaria, säde-eliöt
- Foraminifera, huokoseliöt
- Cercozoa
- Archaeplastida (mm. kasvit)
- Unikonta (yksisiimaiset, mm. eläimet)

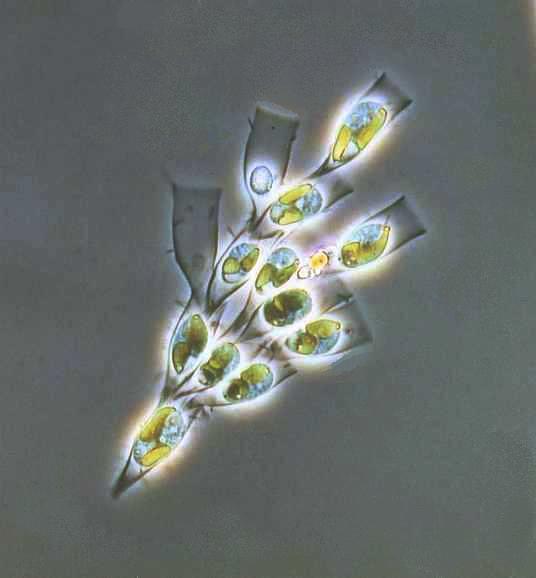
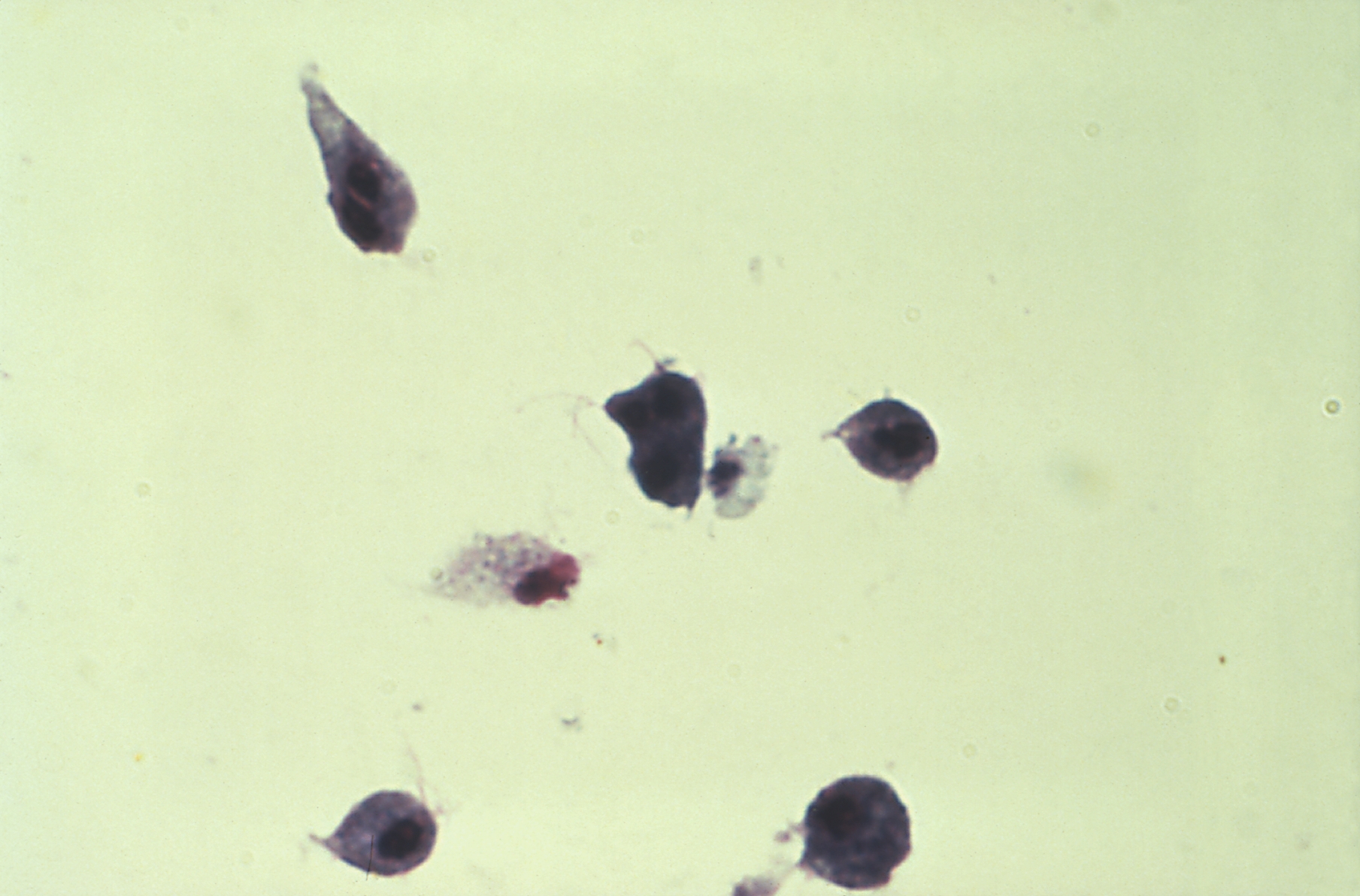
Huokoseliöitäkin (Foraminifera) pidettiin aiemmin yksisoluisina alkueläiminä; siitä nimitys huokoseläimet. Niiden muodostaman pääjakson sekä suomalainen että tieteellinen nimi johtuu niiden soluruumista peittävästä kuoresta (testa), joka on täynnä huokosia (lat. foramen; ferre ’kantaa, kuljettaa’). Kuori on yksiosainen, mutta usein monilokeroinen, ja kalsiumkarbonaatin vahvistama. Outoa kyllä yksisoluisten kyseessä ollen, mutta suurimmat huokoseläinkuoret ovat halkaisijaltaan useita senttimetrejä.
Valejalat pistävät ulos kuoren huokosista ja toimivat uintieliminä, kuoren rakentajina ja ravinnon hankkijoina. Monet huokoseliöt saavat ravintoa myös symbionttisten levien fotosynteesistä.
Huokoseliöitä tavataan sekä merissä että sisävesissä. Enin osa elää hiekanjyvien seassa tai kiinnittyneenä kiviin tai leviin. On myös planktisia lajeja.
Kaikista tunnetuista huokoseliölajeista 90 % on fossiilisia. Yhdessä muiden kalkkipitoisten eliöiden kanssa huokoseliöt muodostavat merien pohjaan kerrostumia. Muinaisista pohjakerrostumista on muodostunut myös kallioita, jotka nykyään ovat kuivaa maata. Niiden ikää voidaan päätellä niiden sisältämistä huokoseliöfossiileista, ja eri puolilla maapalloa olevat muodostumat voidaan osoittaa samanikäisiksi. Ne paljastavat myös ilmastossa tapahtuneita muutoksia ja niiden vaikutuksia meriin ja niiden elämään.
Erään viimeaikaisen luokittelun mukaan (Mikhalevich 2013) huokoseliöiden pääjakso käsittää kuusi luokkaa, joista useimmissa on useita lahkoja. Lahko Reticulomyxida on systemaattiselta asemaltaan epävarma, eikä sitä ole kyetty sijoittamaan mihinkään luokkaan. Edellä mainittu luokittelu käsittää seuraavat luokat ja lahkot (alaluokat, ylälahkot ym:t välikategoriat on tässä jätetty huomiotta):
Luokka 1. Schizocladea, ainoa lahko Schizocladida.
Luokka 2. Xenophyophorea. Tärkeitä merien pohjaeliöitä, joita tavataan kaikissa valtamerissä mannerjalustoilta syvänteisiin; solu monitumainen; kuori kooltaan muutamasta millimetristä jopa 20 senttimetriin, muodoltaan vaihteleva, joskus rakenteeltaan monimutkainen; monimuotoinen luokka. Kaksi lahkoa: Stannomida ja Psamminida. – kuvia
Luokka 3. Astrorhizata. Kuori yksilokeroinen ja koostuu yhteen liimautuneesta materiaalista tai kalkista; muodoltaan pallomaisia, pitkänomaisia tai epäsäännöllisiä. Kahdeksan lahkoa: Ammoscalariida, Lagynida, Allogromiida, Astrorhisida, Dendrophryida, Hippocrepinida, Parathuramminida (kuollut sukupuuttoon), Psammosphaerida. – Lahkon Allogrimiida lajien kuori on enimmäkseen orgaanista ainesta, muodoltaan yksinkertainen, yksikammioinen; sekä mereisiä että sisävesilajeja; varhaisimmat huokoseliöfossiilit kuuluvat tähän lahkoon. – kuvia
Luokka 4. Rotaliata. Kuori moniosainen, enimmäkseen spiraalimainen; siinä yksi tai useampia aukkoja; kuoren sisäosa voi olla monimutkainen sekundaarisine kammioineen ja niitä yhdistävine kanavineen; kuori koostuu lasimaisesta kalsiitista tai yhteen liimautuneesta materiaalista. 24 lahkoa: Cassigerinellida, Globigerinida, Hantkenidia, Heterochelicida, Globorotaliida, Nautiloculinida, Spiroplectamminida, Textulariida, Trochamminida, Verneulinida, Robertinida, Elphidiida, Nummulitida, Orbitoidida (sukupuuttoon kuollut), Nonionida, Cassidulinida, Buliminida, Bolivinitida, Chilostomellida, Discorbida, Glabratellida, Planorbulinida, Rotaliida ja Rosalinida. – kuvia
Lahko Globigerinida. Merten planktonissa; kuori lasimaista kalsiittia, muodoltaan spiraalimainen, pitkäpiikkinen; kuori muodostaa pallomaisia kammioita ja tavallisesti siinä on yksi aukko, joskus useampia; fossiileja jurakaudelta lähtien; lajeja tunnetaan yhteensä yli 400, joista nykyään eläviä noin 30; tunneuin suku on Globigerina, jonka aiheuttamaa liejua on laajoilla alueilla merien pohjalla. – kuvia
Lahko Textulariida. Kuori muodostuu yhteen liimautuneesta ja kalkkisementin sitomasta materiaalista; fossiileja kambrikaudelta lähtien. – kuvia
Lahko Trochamminida. Fossiileja kambrikaudelta lähtien. – kuvia
Lahko Robertinida. Fossiileja keski-triaskaudelta lähtien; merten pohjaeliöitä; kuori spiraalimainen, monikammioinen, lasimainen. – kuvia
Lahko Buliminida. Enimmäkseen merenpohjalla; kuori huokoista, lasimaista kalsiittia; kammiot kaden tai kolmen sarjoissa ainakin varhaisvaiheessa. – kuvia
Lahko Rotaliida. Enimmäkseen valtamerten pohjaeliöstössä, eräät lajit jokisuistoissa; kuori lasimaista kalsiittia, monilokeroinen, tyypillisesti kierteinen; kammiot voivat olla sekundaaristen väliseinien jakamia; kuoren pinta voi olla eri tavoin nystyinen tai harjuinen; aukko samoin varustukseltaan vaihteleva. Lahkoon kuuluu paljon tärkeitä fossiileja, kuten nummuliitit. – kuvia
Luokka 5. Nodosariata. Lahkot Ammomarginulinida, Nouriida, Pseudopalmulida (sukupuuttoon kuollut), Saccamminida, Hormosinida, Biseriamminida (sukupuuttoon kuollut), Delosinida, Lagenida, Palaeotextulariida (hävinnyt), Polymorphinida, Vaginulinida ja Nodosarida. – kuvia
Lahko Lagenida. Merten pohjaeliöstöä; kuori kalsiittia, kammiot peräkkäin 1-3 sarjassa; kuoren muodot eräillä lajeilla viuhkamaisia tai lehtimäisiä; fossiileja ylä-siluurikaudelta lähtien. – kuvia
Luokka 6. Spirillinata. Spiraalimaisia, kuori joko yhteen liimautunutta materiaalia tai kalkkia. Lahkot: Plagioraphida (sukupuuttoon kuollut), Ammodiscida, Ammovertellinida, Ataxophragmiida, Archaediscida (sukupuuttoon kuollut), Lasiodiscida (sukupuuttoon kuollut), Tetradaxida (sukupuuttoon kuollut), Hottingerellida (sukupuuttoon kuollut), Involutinida (fossiileja varhais-permikaudelta myöhäis-liitukaudelle), Seabrookiida, Cymbaloporida, Spirillinida (kuori putkimainen ja kiertynyt, kalsiitista muodostunut; sukusolut eli gameetit ovat amebamaisia, kun taas muilla huokoseliöillä ne ovat kaksisiimaisia), Patellinida, Miliolata (muodoltaan vaihtelevia; kuori posliinimainen, joillakin siihen liittyy ympäristöstä peräisin olevia aineksia), Lituotubida, Loftusiida, Sphaeramminida, Cyclolinida, Haplophragmiida, Schlumbergerinida (eli Rzehakinida), Lituolida (fossiileja kambrikaudelta alkaen), Ozawainellida (sukupuuttoon kuollut), Endothyroida (sukupuuttoon kuollut), Tournayellida (sukupuuttoon kuollut), Fusulinida (sukupuuttoon kuollut; ala-siluurikaudelta ylä-permikaudelle; lajit ovat erinomaisia johtofossiileja kerrostumien ikää määritettäessä; paikoin niiden kuoret ovat merkittävä osa kalkkikiveä), Neoschwagerinida (sukupuuttoon kuollut), Schubertellida (sukupuuttoon kuollut), Schwagerinida (sukupuuttoon kuollut), Staffellida (sukupuuttoon kuollut), Costiferida (sukupuuttoon kuollut), Squamulinida, Cornuspirida, Soritida, Nebeculariida, Miliolida (kalkkikuori posliinimainen, sisäosa tavallisesti pseudokitiiniä; kuori magnesiumpitoinen ja orgaanista ainesta sisältävä, huokoseton ja monikammioinen; fosiileja kivihiilikaudelta alkaen; merten pohjaeliöstöä rannikoilta syvänteisiin). – kuvia




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}