Hiekkaisilta tai mutaisilta merten pohjilta löytyy pieniä, pehmeäruumiisia eläimiä, jotka elävät yksinään tai ryhmissä, ja joita nimitetään esiselkäjänteisiksi (pääjakso Hemichordata). Niiden ruumis ja ruumiinontelo jakautuvat kolmeen osastoon, niillä on tavallisesti parilliset kidusaukot, ja hermokudosta on sekä selkä- että vatsapuolen ihossa. Suuontelosta kohti eläimen etupäätä lähtee taskumainen muodostuma kudosta, joka muistuttaa selkäjänteisten (Chordata) selkäjännettä. Näillä kahdella pääjaksolla on muitakin yhteisiä rakennepiirteitä.
Esiselkäjänteiset ja selkäjänteiset muistuttavat toisiaan nieluun liittyvien kidusaukkojen osalta. Edellisten etupäässä on hermoston osa, joka alkionkehityksessä saa alkunsa uloimmasta solukerroksesta (epidermistä), on toisinaan ontto ja sijaitsee keskiruumiin selkäpuolella. Näin ollen se muistuttaa selkäjänteisten keskushermostoa. Alkio ja varhainen toukkavaihe taas muistuttavat meritähtien vastaavia: molemmilla on ripsivyöhykkeet, samankaltainen ruoansulatuskanava sekä erityisesti ruumiinontelon jakautuminen kolmeksi osastoksi. Nämä piirteet viittaavat vahvasti siihen, että kaikilla kolmella – piikkinahkaisilla (joihin meritähdet kuuluvat), esiselkäjänteisillä ja selkäjänteisillä – on yhteinen kantamuoto.
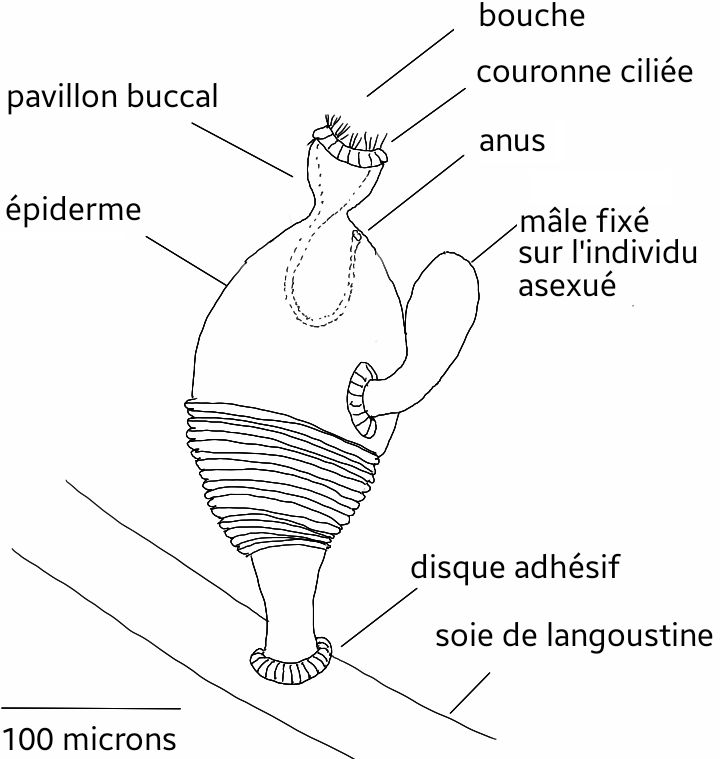
Esiselkäjänteisten ruumiissa on päältäpäinkin erotettavissa kolme osaa: proto-, meso- ja metasomi eli etupää, keskiruumis ja takaruumis. Silti ruumiinmuoto voi olla aika erilainen: toiset ovat matomaisia (ns. terhomadot), toiset pikarimaisia (ns. siipikiduksiset). Jo puheena olleita parillisia kidusaukkoja on monta, kaksi tai ne puuttuvat. Ruumiin seinämä muodostuu ihosta eli epidermistä, hermokudoksesta, lihassyistä ja ruumiinontelon kalvosta eli peritoneumista.
Sisäelimiin kuuluu em. eturuumiin suuontelotaskun lisäksi suora tai U:n muotoinen ruoansulatuskanava. Edellisessä tapauksessa peräaukko on takaruumiin päässä, jälkimmäisessä suun lähellä. Ruumiinontelon kolmesta osastosta keski- ja takaruumiin ontelot ovat parilliset. Verenkierto on avoin; keskiruumiin selkä- ja vatsapuolella on suonet ja eturuumiissa sijaitsee sykkivä suoni. Hermosto on hajanainen, se sijaitsee ihossa eläimen selkä- ja vatsapuolen keskellä.
Lisääntymiseltään esiselkäjänteiset ovat yksineuvoisia, mikä merkitsee, että sukupuolet ovat erilliset. Ulkonaisesti sukupuolia ei voi erottaa toisistaan. Terhomadoilla on monta parillista sukurauhasta. Toukkavaihetta nimitetään tornariaksi, mutta se voi puuttuakin. Siipikiduksisilla on kaksi sukurauhasta ja kehityksessään toukkavaihe. Aikuiset voivat lisääntyä suvuttomasti silmikoimalla.
Esiselkäjänteisten pääjakso, Hemichordata, käsittää yli 120 nykyistä lajia ja jaetaan kolmeksi luokaksi:
- Terhomadot, Enteropneusta: matomaisia, yksinään eläviä, pituus 2,5 – 25 cm, pehmeitä, kokoon vetäytyviä; ruumis jakautunut kärsäksi, kaulukseksi ja takaruumiiksi; parillisia kidusrakoja 10 – 80 tai jopa enemmän; suoli suora; ruumiinontelo selvästi erottuva ja 3-osainen; lonkerottomia. Sukuja: Ptychodera, Balanoglossus, Saccoglossus.
- Siipikiduksiset, Pterobranchia. Enintään 7 mm pitkiä, ruumis pikarimainen; elävät tavallisesti yhdyskuntina, jotka eläinten erittämän kuoren suojassa; suoli U:n muotoinen; kidusaukkoja 2 tai ne puuttuvat; kauluksessa lonkerokannatin (lofofori), jossa lonkeroita; lisääntyvät suvullisesti tai silmikoimalla. Luokka sisältää ilmeisesti myös graptoliitit (Graptolithina), jotka elivät merissä kelluvia yhdyskuntia muodostaen keskikambrikaudelta hiilikauden alkuvaiheisiin.
- Luokassa Planctosphaeroidea on vain yksi laji, josta tunnetaan vain toukkavaihe.
Erityisesti merten syvänteistä on löydettävissä entuudestaan tuntemattomia lajeja.
Esiselkäjänteiset muodostavat piikkinahkaisten ja selkäjänteisten kanssa uusi- eli jälkisuisten pääjaksoryhmän (Protostomia), joka on saanut nimensä alkionkehityksessä tapahtuvan alkusuun muodostumisesta peräaukoksi ja suun muodostumisesta uudestaan.